
【文獻(xiàn)解讀】HIC揭秘飲食誘發(fā)肥胖過程中啟動子錨定的染色質(zhì)互作動力學(xué)
信息來源:金開瑞 作者:genecreate 發(fā)布時間:2020-06-01 16:24:19
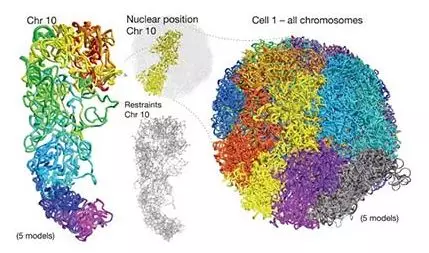
關(guān)于Hi-C
Hi-C(High-through chromosome conformation capture)技術(shù)源于染色體構(gòu)象捕獲 (Chromosome conformation capture, 3C)技術(shù),以整個細(xì)胞核為研究對象,利用高通量測序技術(shù),結(jié)合生物信息分析方法,研究全基因組范圍內(nèi)整個染色質(zhì)DNA在空間位置上的關(guān)系,通過對染色質(zhì)內(nèi)全部DNA相互作用模式進(jìn)行捕獲,獲得高分辨率的染色質(zhì)三維結(jié)構(gòu)信息。
(引自Pueschel R, Coraggio F, et al. From single genes to entire genomes: the search for a function of nuclear organization. Development , 2016 , 143 (6) :910)
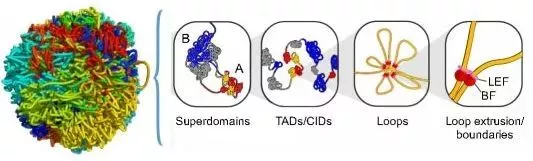
通過Hi-C技術(shù)可以獲得全基因組范圍內(nèi)的互作信息,得到染色體三個層級的三維結(jié)構(gòu):A/B compartment、拓?fù)湎嚓P(guān)結(jié)構(gòu)域(TAD)、染色質(zhì)環(huán)(loop)。
解析文獻(xiàn)
Alterations in promoter interaction landscape and transcriptional network underlying metabolic adaptation to diet
期刊:Nature Communications
IF:11.878
背景
為應(yīng)對可能對健康產(chǎn)生重大影響的食物供應(yīng)和食物營養(yǎng)含量的波動,動物對營養(yǎng)質(zhì)量和有效性的響應(yīng)范圍已經(jīng)進(jìn)化到從完全缺乏營養(yǎng)素到過量。隨著西方生活方式的普及,高脂肪、高能量飲食,肥胖和NAFLD成為人們?nèi)找骊P(guān)注的健康問題。在卡路里過多的情況下,肝臟通過動態(tài)轉(zhuǎn)錄組重編輯來適應(yīng)營養(yǎng)過剩,這是維持代謝體內(nèi)平衡所必需的多種適應(yīng)之一。目前基因調(diào)控對介導(dǎo)肝臟代謝適應(yīng)過多卡路里攝入的機制未完全了解,對啟動子-增強子互作動力學(xué)的詳細(xì)分析,可以了解從營養(yǎng)誘導(dǎo)的代謝適應(yīng)到疾病發(fā)展的過程。增強子通常距離其靶基因很遠(yuǎn),因此難以準(zhǔn)確地研究將特定的增強子分配給其靶基因及基因調(diào)控的細(xì)節(jié)。Hi-C被應(yīng)用于識別全基因組染色質(zhì)相互作用,并提供對高階染色質(zhì)組織的了解。
作者采用飲食誘導(dǎo)肥胖的C57BL / 6J小鼠模型,在這些小鼠的肝臟組織上進(jìn)行了Hi-C和PCHi-C,結(jié)合ChIP-seq和RNA-seq,以研究飲食誘發(fā)肥胖過程中啟動子錨定的染色質(zhì)互作動力學(xué)。
結(jié)果
1、慢性肥胖致肝臟轉(zhuǎn)錄組重編
作者對雄性C57BL / 6小鼠進(jìn)行了飲食研究,五周大的小鼠適應(yīng)普通飲食(NIH-31),隨后(每組5只小鼠)接受高碳水化合物和高脂的飲食20周。在研究過程中,高脂飲食的小鼠明顯肥胖,在20周后總體重幾乎是高碳水化合物飲食的兩倍。肥胖小鼠的葡萄糖和胰島素耐受性較差,血漿胰島素和瘦蛋白水平顯著增加,這表明它們的代謝功能異常。
為了確定肥胖如何影響全身代謝,將喂食LD(高碳水化合物飲食)和CD(高脂飲食)的小鼠測量其耗氧量(VO 2),二氧化碳產(chǎn)生量(VCO 2)和能量消耗(EE)。與CD組相比,LD組的平均EE,VO 2和VCO 2值降低,血漿總膽固醇,高密度脂蛋白(HDL),低密度脂蛋白(LDL),天冬氨酸轉(zhuǎn)氨酶(AST)和丙氨酸轉(zhuǎn)氨酶(ALT)明顯升高。LD組脂肪組織和肝臟質(zhì)量均顯著高于CD組,脂肪細(xì)胞大小增加。
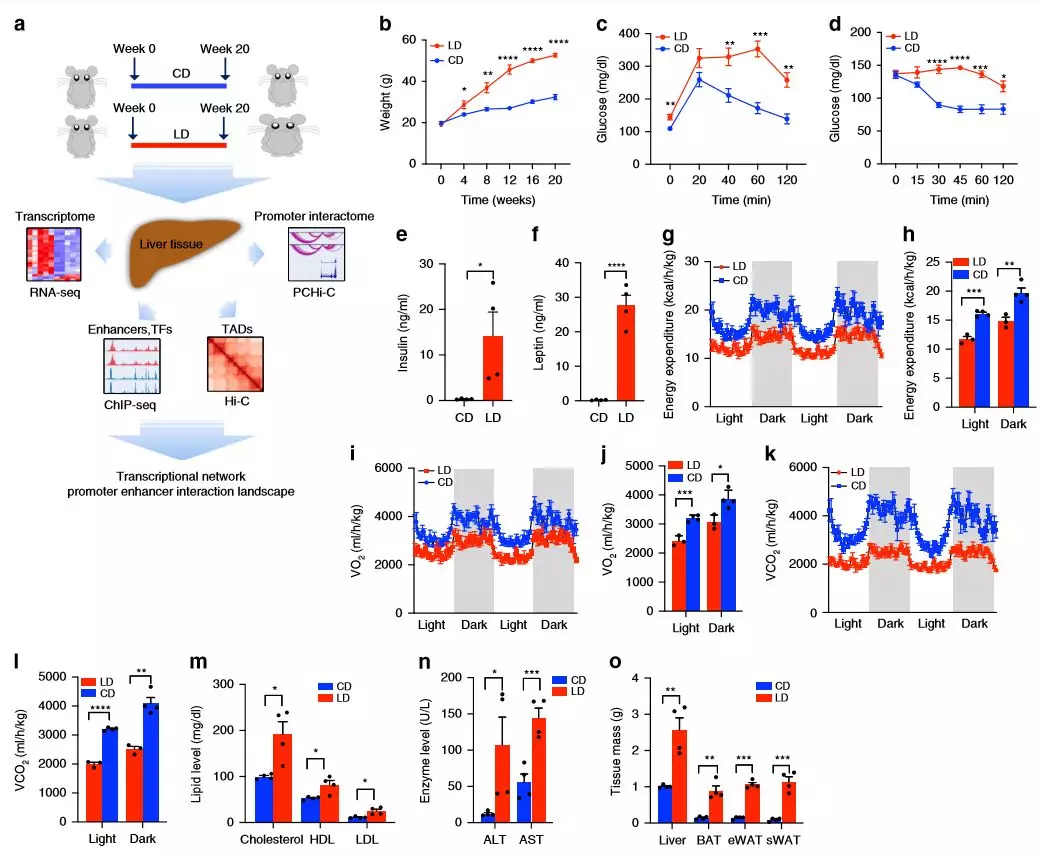
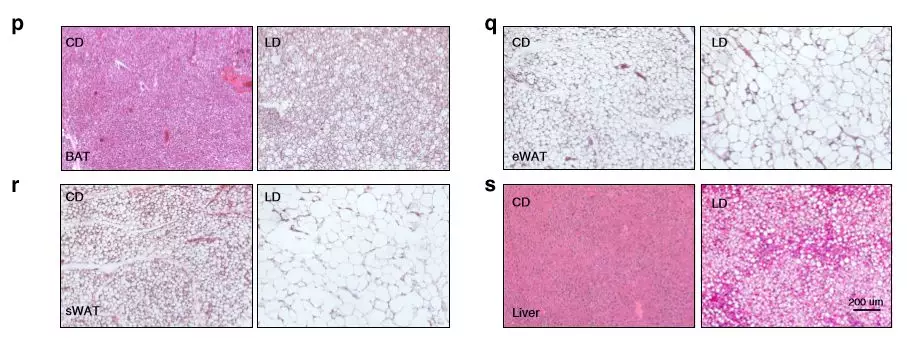
圖1飲食對代謝參數(shù)的影響
為了解肥胖的轉(zhuǎn)錄反應(yīng),作者對研究小鼠(每組5只)的肝臟進(jìn)行RNA-seq,差異表達(dá)基因(DEG)分析確定了高脂肪飲食的肥胖組中有2066個基因上調(diào),而高碳水化合物飲食組有1663個基因上調(diào)。在慢性肥胖和NAFLD中,脂質(zhì)代謝受到干擾,其特征是介導(dǎo)新脂質(zhì)生成(DNL)的基因下調(diào),而涉及脂肪酸氧化的基因(FAO)上調(diào)。轉(zhuǎn)錄組中的差異反映了高脂質(zhì)和高碳水化合物的飲食在小鼠肝臟中活躍的代謝狀態(tài)和調(diào)控網(wǎng)絡(luò)的差異。
2、慢性肥胖改變了肝臟的增強子網(wǎng)絡(luò)
為了解適應(yīng)肥胖是如何影響增強子活性,作者通過肝臟組織ChIP-seq對H3K27ac(每組3只)進(jìn)行了分析。研究發(fā)現(xiàn)差異富集的基因座大多位于遠(yuǎn)離TSS的位置,并且被歸類為增強子。高脂飲食組中富含H3K27ac的區(qū)域,最富集的基序?qū)?yīng)于ETS(ETS1,EHF),bZIP(FOSL2,JUN-AP1和ATF3)和C / EBP(C / EBPA, C / EBPB和C / EBPE)家族轉(zhuǎn)錄因子。
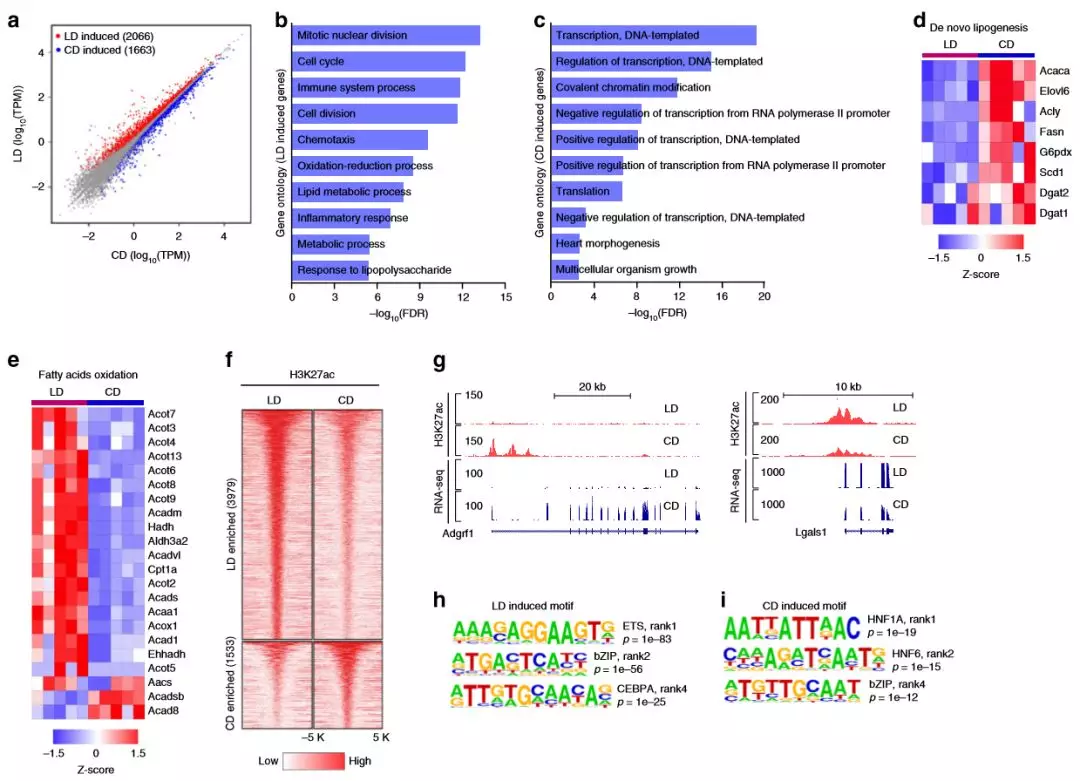
圖2慢性肥胖的肝臟轉(zhuǎn)錄組和增強子網(wǎng)絡(luò)會重編輯
3、肥胖不會改變與拓?fù)湎嚓P(guān)結(jié)構(gòu)域
為詳細(xì)了解染色質(zhì)互作,作者在高碳水化合物和脂質(zhì)的飲食中對小鼠(每組2只)的肝臟進(jìn)行了原位Hi-C處理。與預(yù)期一樣,飲食和肥胖及其所有相關(guān)的生理和代謝變化均未影響A和B區(qū)室的模式。作者比較了兩組之間的TAD邊界,約90%的邊界相同,表明在肥胖和NAFLD期間高階染色質(zhì)組織保持不變。
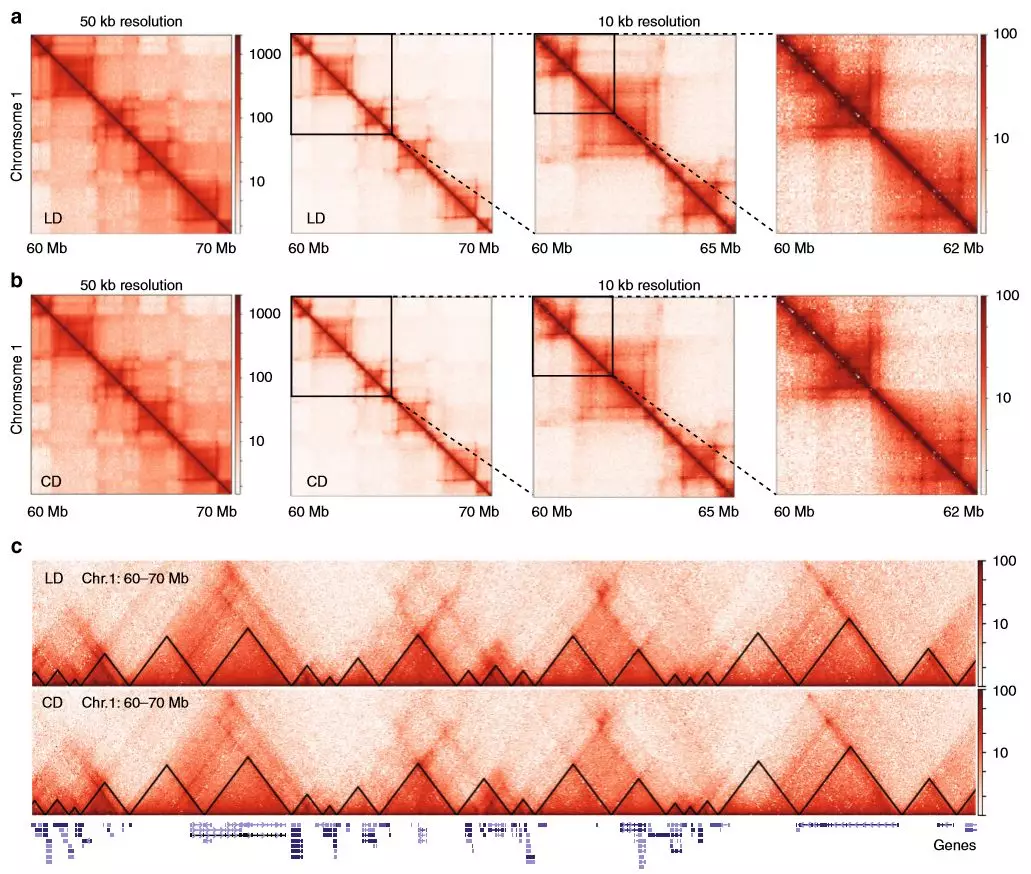
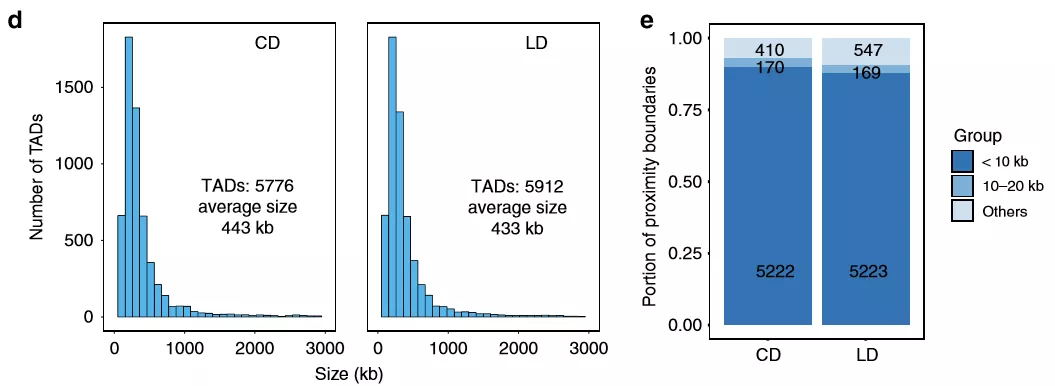
圖3慢性肥胖肝臟中的高級染色質(zhì)結(jié)構(gòu)
4、在代謝適應(yīng)中促進(jìn)染色質(zhì)相互作用
作者在Hi-C數(shù)據(jù)中發(fā)現(xiàn)高脂質(zhì)飲食的肝臟中34,982個顯著的染色質(zhì)互作,以及高碳水化合物飲食中37,185個顯著的染色質(zhì)互作,分別有28,682和30,502個染色質(zhì)相互作用與基因啟動子(TSS +/- 2 kb)重疊。為獲得肝臟中啟動子錨定的染色質(zhì)互作及其在適應(yīng)肥胖后的變化的高分辨率視圖,作者進(jìn)行了啟動子捕獲Hi-C(PCHi-C)(每組2只)。LD和CD組共享了很大一部分交互作用(54%),這表明許多啟動子互作是穩(wěn)定的。幾乎80%的相互作用跨越<500 kb,大約13%的相互作用發(fā)生在兩個啟動子之間,表明啟動子調(diào)控遠(yuǎn)端基因元件。
為了解啟動子相互作用對基因表達(dá)的潛在調(diào)控作用,作者將啟動子互作與基因表達(dá)相關(guān)聯(lián)。通常,啟動子互作的數(shù)量增加與基因表達(dá)呈正相關(guān),作者還觀察到也富集了未表達(dá)基因的啟動子互作,這表明某些啟動子相互作用是預(yù)先形成的,而不是由增強子激活誘導(dǎo)的。作者測試了H3K27ac富集與啟動子互作區(qū)域(PIR)的交集,值得注意的是,具有H3K27ac富集的PIR與基因表達(dá)的增加顯著相關(guān),這支持啟動子與活性增強子的接觸會上調(diào)基因表達(dá)的模型。
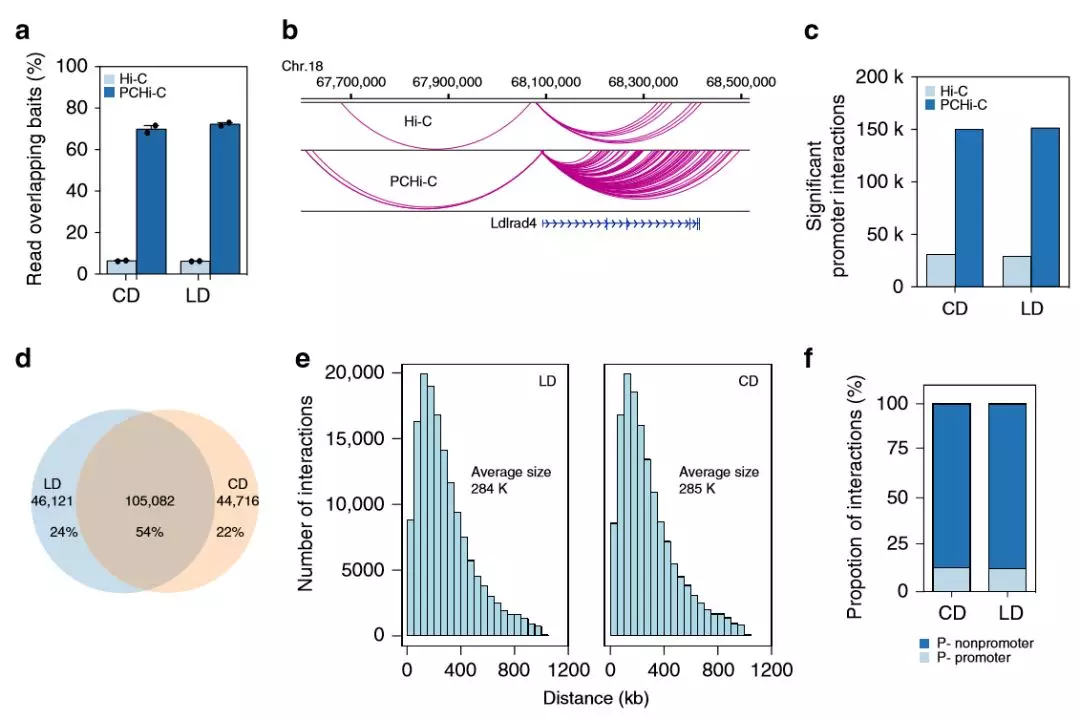
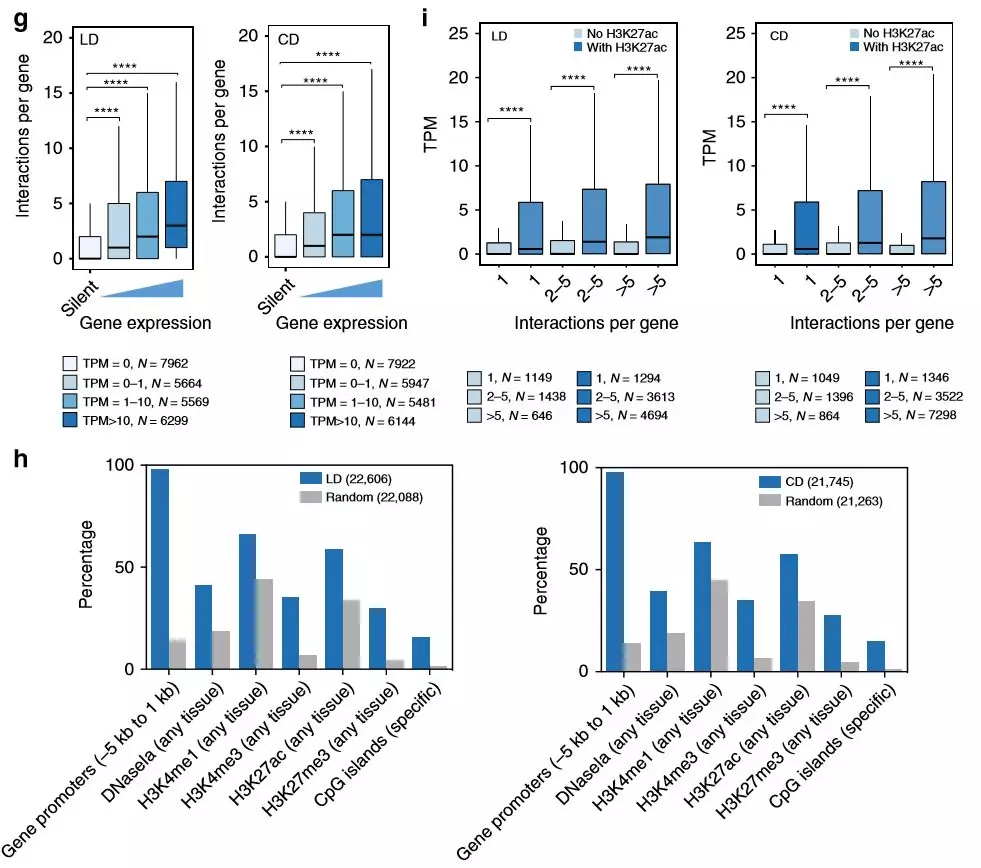
圖4在代謝適應(yīng)飲食中啟動子錨定的染色質(zhì)互作調(diào)節(jié)的基因表達(dá)變化
5、差異表達(dá)基因的啟動子接觸改變
為了解代謝適應(yīng)飲食是否改變了肝臟中的啟動子環(huán)化,作者使用了edgeR 來鑒定染色質(zhì)相互作用模式的差異。其中1962個重編的啟動子相互作用中,高脂質(zhì)的飲食中增加705次,而高碳水化合物的組中增加1257次,分別與109和228 個DEG重疊。比較與靜態(tài)環(huán)相關(guān)的DEG,只有對高碳水化合物的飲食有反應(yīng)的啟動子/增強子相互作用與基因表達(dá)的增加顯著相關(guān)。這表明,新形成的啟動子互作有助于代謝基因的轉(zhuǎn)錄調(diào)控,并且可能與飲食有關(guān)。
6、預(yù)形成的啟動子-增強子相互作用
為了解在Hi-C數(shù)據(jù)中,與啟動子互作的增強子的活性狀態(tài)是否反映了基因表達(dá),作者檢查了差異富集的H3K27ac與啟動子遠(yuǎn)端互作位置的重疊,發(fā)現(xiàn)了4449個啟動子-增強子相互作用,其中H3K27ac在高脂質(zhì)的飲食組中差異性富集。無論飲食如何,啟動子與乙酰化增強子互作的DEG均上調(diào),在預(yù)形成的環(huán)中,增強子的激活狀態(tài)發(fā)生變化的啟動子-增強子環(huán)的數(shù)量(6452)大大超過了條件變化后形成的新環(huán)的數(shù)量(1962)。這些發(fā)現(xiàn)表明,已經(jīng)與啟動子互作的增強子的激活,代表了代謝應(yīng)激期間遠(yuǎn)端基因調(diào)控的一種重要模式。
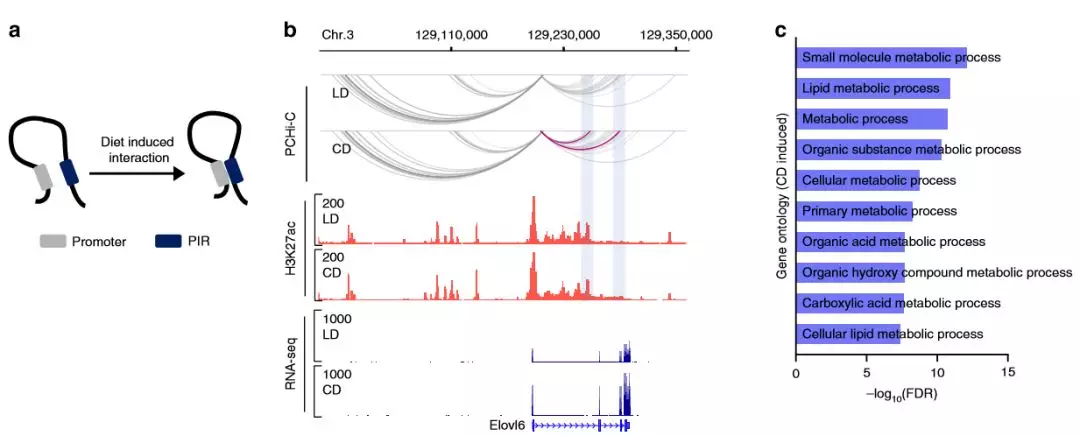
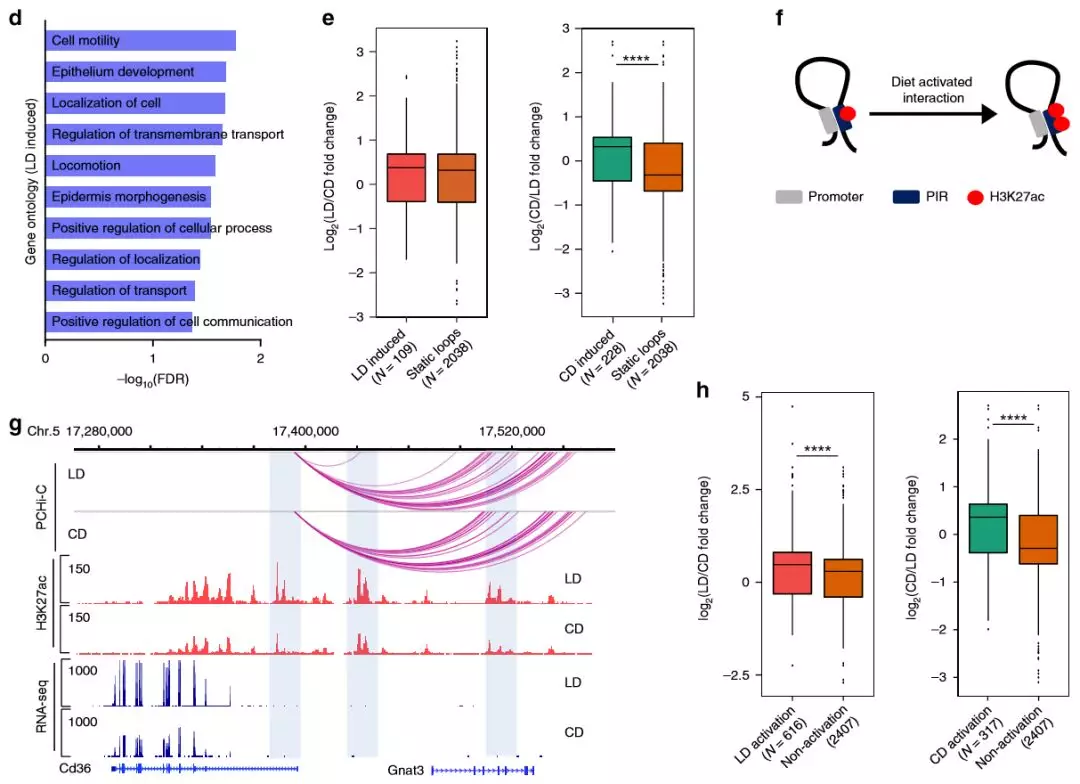
圖5 飲食改變了啟動子互作
7、轉(zhuǎn)錄組分析將Hnf4α與代謝聯(lián)系起來
為了評估哪些轉(zhuǎn)錄因子結(jié)合參與基因表達(dá)變化,作者在啟動子中進(jìn)行了基序分析,觀察到包括Hnf4α 的幾個TF基序的富集 ,HNF4α是信號響應(yīng)轉(zhuǎn)錄因子,在調(diào)節(jié)脂質(zhì)代謝中起重要作用。作者通過qPCR和免疫印跡,發(fā)現(xiàn)肥胖組中Hnf4α在蛋白質(zhì)和mRNA水平上的下調(diào),C18多不飽和脂肪酸包括亞油酸是HNF4α的已知配體。接下來,作者進(jìn)行了ChIP-seq(每組2只),以了解Hnf4α如何響應(yīng)肥胖。肥胖組中更多的Hnf4α富集峰與脂質(zhì)配體對Hnf4α的活化相一致,作者還觀察到啟動子的遠(yuǎn)端互作與Hnf4α結(jié)合重疊,這表明Hnf4α與啟動子互作的位點結(jié)合。啟動子遠(yuǎn)端互作位點Hnf4α峰增加,DEGs也顯示基因表達(dá)上調(diào)。
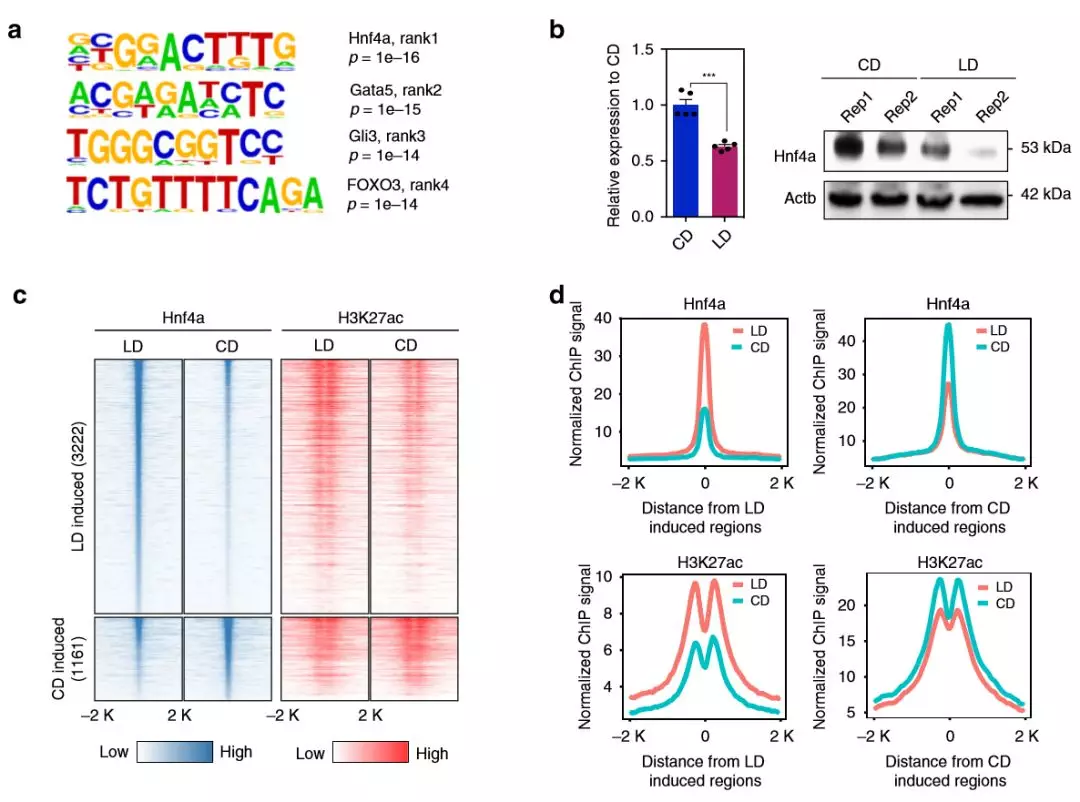
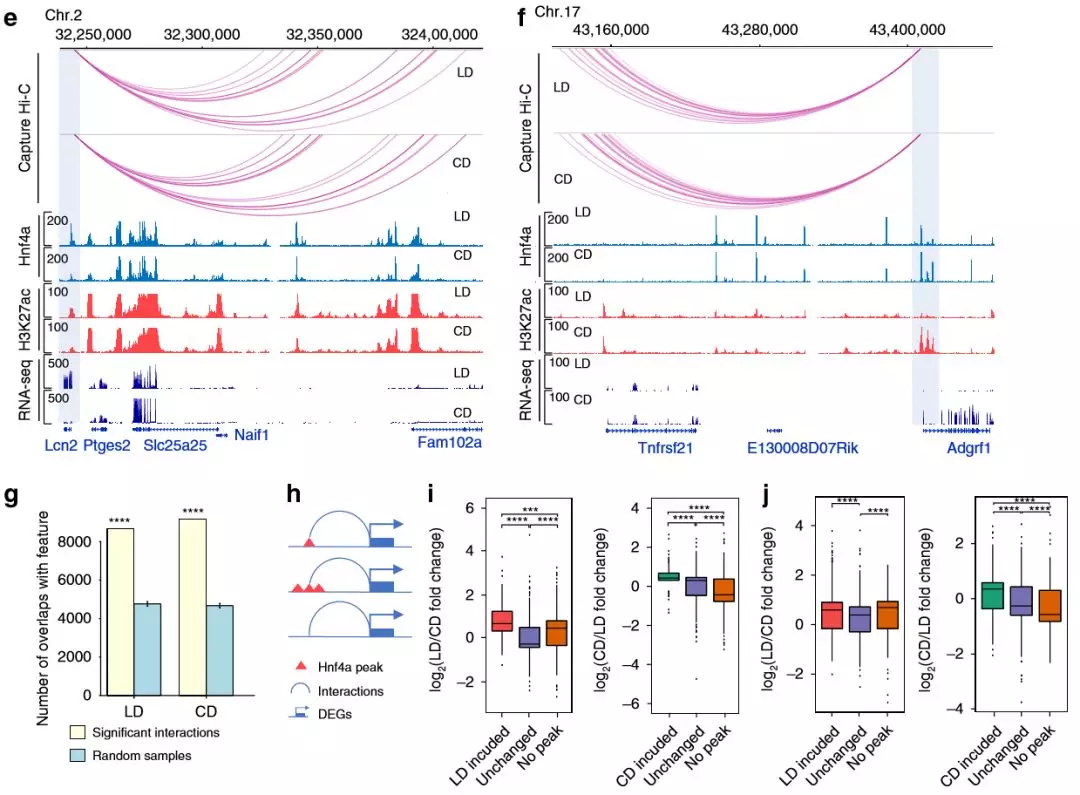
圖6 轉(zhuǎn)錄組分析將Hnf4α與NAFLD的代謝基因的調(diào)節(jié)聯(lián)系起來
8、Hnf4α與其他轉(zhuǎn)錄因子結(jié)合
盡管作者確定了大約4000個發(fā)生了變化的Hnf4α結(jié)合位點,但這個數(shù)字僅代表約10%的Hnf4α的全基因組結(jié)合位點。在所有Hnf4α結(jié)合位點中,有79%與富集H3K27ac的基因座共定位,這表明肝臟中大多數(shù)已鑒定的Hnf4α結(jié)合區(qū)域是在兩種情況下都存在且活性元件或開放染色質(zhì)不易改變。如此大比例的Hnf4α結(jié)合位點是穩(wěn)定的,作者假設(shè)Hnf4α可能在代謝應(yīng)激期間與其他轉(zhuǎn)錄因子發(fā)生功能性相互作用,研究了Hnf4α結(jié)合的激活區(qū)域中存在哪些轉(zhuǎn)錄因子結(jié)合基序,最顯著富集的共有基序是HNF4α,第二個轉(zhuǎn)錄因子是基本亮氨酸拉鏈結(jié)構(gòu)域(bZIP)家族,其中包括C / EBP家族。
為了驗證基序結(jié)果,作者進(jìn)行了C /EBPαChIP-seq(每組2只),發(fā)現(xiàn)在肝臟中有55%的C /EBPα峰與Hnf4α共定位,共結(jié)合位點涉及多種代謝途徑的基因,包括羧酸代謝,脂肪酸代謝,類固醇代謝和脂質(zhì)分解代謝。作者在C /EBPα差異結(jié)合位點檢查了Hnf4α結(jié)合信號,Hnf4α信號在差異富集的C /EBPα區(qū)域較高,表明至少在這些位點上始終存在Hnf4α。這些數(shù)據(jù)表明,在與飲食的代謝適應(yīng)過程中,Hnf4α經(jīng)常與C /EBPα結(jié)合以調(diào)節(jié)基因表達(dá)。
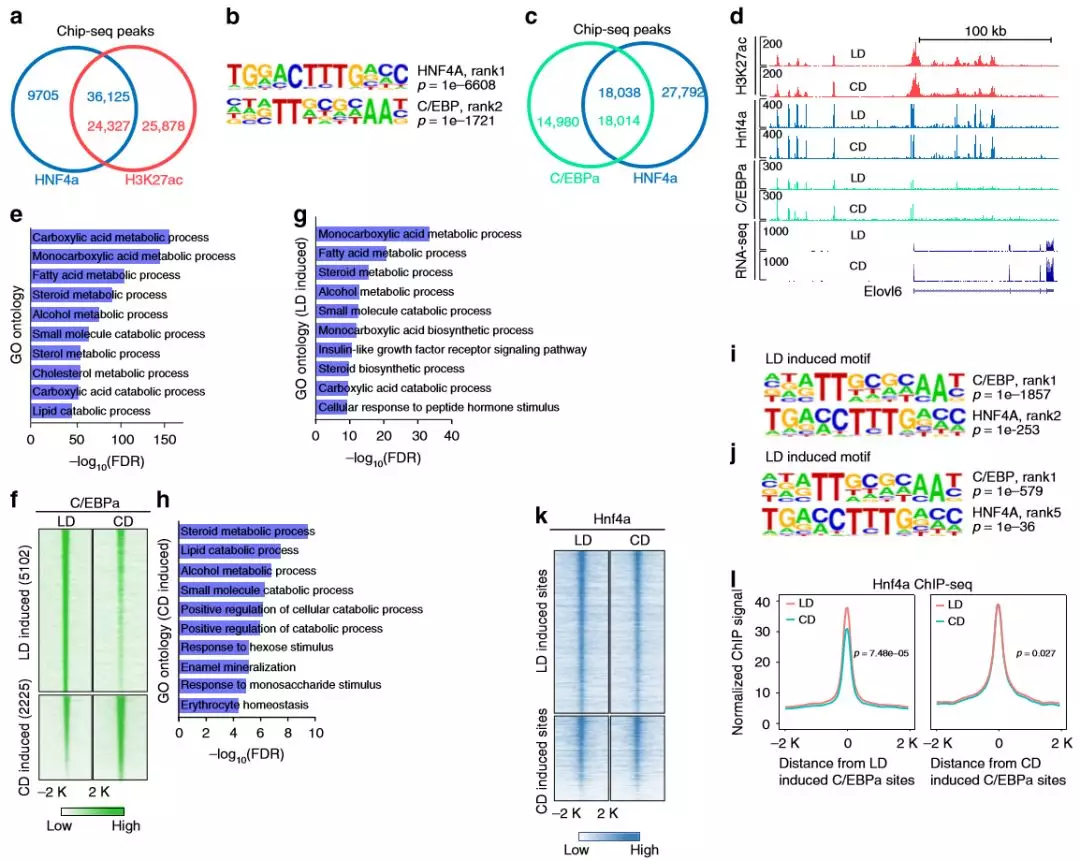
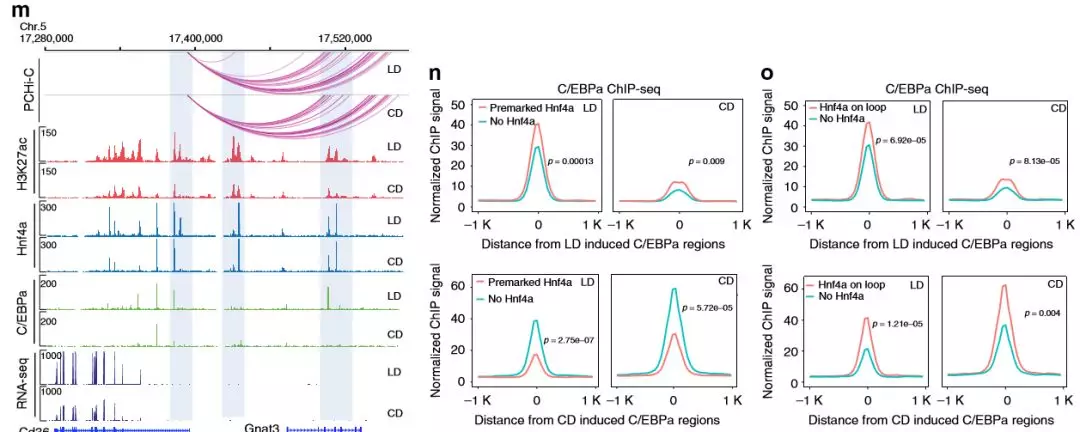
圖7 Hnf4α經(jīng)常與C /EBPα共結(jié)合
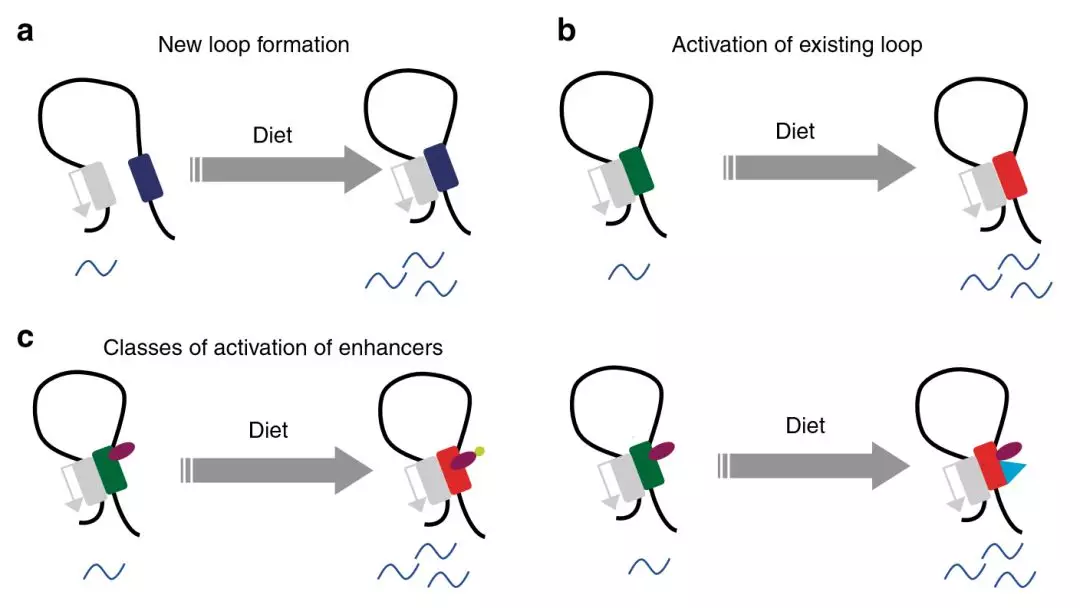
圖8代謝適應(yīng)飲食的啟動子/增強子互作
總結(jié)
代謝適應(yīng)營養(yǎng)狀態(tài)需要改變關(guān)鍵組織中的基因表達(dá)。作者研究了染色質(zhì)相互作用動力學(xué),以及在小鼠模型系統(tǒng)中順式調(diào)控基因座和轉(zhuǎn)錄網(wǎng)絡(luò)的變化。與高碳水化合物的飲食相比,長期攝入高脂肪的飲食會導(dǎo)致肝臟轉(zhuǎn)錄網(wǎng)絡(luò)發(fā)生嚴(yán)重的重編。通過啟動子捕獲Hi-C技術(shù)發(fā)現(xiàn),啟動子與遠(yuǎn)端調(diào)節(jié)基因座的遠(yuǎn)距離相互作用受代謝狀態(tài)的調(diào)節(jié),而代謝狀態(tài)具體取決于飲食。對高脂飲食的適應(yīng)主要由包括Hnf4α在內(nèi)的受體介導(dǎo),這適應(yīng)取決于增強子/啟動子環(huán)激活。適應(yīng)富含碳水化合物的飲食,導(dǎo)致預(yù)先形成環(huán)的活化和新形成的啟動子/增強子互作。這些結(jié)果表明,對營養(yǎng)變化和代謝應(yīng)激的適應(yīng)性是通過從頭和已有的染色質(zhì)互作而發(fā)生的,它們對代謝信號的反應(yīng)不同。
畫重點
公眾號:金開瑞生物,對話框回復(fù):飲食誘發(fā)肥胖
即可獲得全文PDF
關(guān)于我們
武漢金開瑞生物工程有限公司提供的DLO Hi-C技術(shù),是一種創(chuàng)新的染色質(zhì)構(gòu)象捕獲技術(shù),此技術(shù)信噪比高,質(zhì)量控制于早期,為解析基因組三維結(jié)構(gòu)提供了一種新型、高效、經(jīng)濟(jì)的研究方案。
技術(shù)優(yōu)勢
1.微量細(xì)胞建庫:正常建match庫與生信分析的樣本量可低至10萬個核。
2.高成功率:細(xì)胞樣本文庫構(gòu)建成功率幾乎為100%。
3.建庫周期短:只需執(zhí)行兩輪簡單的消化和連接步驟即可獲得高質(zhì)量的文庫。
4.數(shù)據(jù)更準(zhǔn)確:測序前質(zhì)檢,確保數(shù)據(jù)準(zhǔn)確性
5.分辨率更高:在測序數(shù)據(jù)量更少的情況下,互作矩陣分辨率更高,染色質(zhì)結(jié)構(gòu)分析得到的數(shù)據(jù)也更多
6. 較高的信噪比:使用多種措施來減少噪音,保證高質(zhì)量的數(shù)據(jù)輸出,分析更準(zhǔn)確。
7. 量身定制個性化分析方案:提供DLO Hi-C的標(biāo)準(zhǔn)分析外,更注重與RNA-Seq、ChIP-Seq、ATAC-Seq和甲基化等多組學(xué)表觀遺傳分析,提供個性化的生信分析方案。
最新動態(tài)
-
09.23
中藥的現(xiàn)代詮釋:外泌體如何革新傳統(tǒng)醫(yī)學(xué)?
-
07.02
1+1>2!深度解析RNA測序數(shù)據(jù)挖掘邏輯和后期實驗設(shè)計思路,輕松研獲10+ SCI
-
07.01
“稻”亦有道——盤點近期水稻研究的重大突破
-
06.28
科學(xué)與美學(xué)的結(jié)合體:植物亞細(xì)胞定位技術(shù)詳解
-
06.28
“聚焦新質(zhì)生產(chǎn)力,激發(fā)科研新動能”|LCA躋身蛋白互作研究的新銳力量
-
06.05
知無不“研”|一文讀懂免疫共沉淀技術(shù)(Co-IP)
-
05.14
四大研究利器(Co-IP、BIFC、Y2H、GST pull-down)助力速配蛋白互作“最佳拍檔”
-
05.14
高效、精準(zhǔn)、直觀、實時——取經(jīng)“蛋白互作研究翹楚”BIFC!
-
05.14
轉(zhuǎn)染效率低、干擾效果差、重復(fù)性欠佳...siRNA研究頻遇“攔路虎”怎么辦?
-
04.22
一文讀懂EMSA技術(shù)核心要點,讓“emsa” 秒變“easy”
 X
X -
 在線客服
在線客服 -

 在線訂購
在線訂購 -

 掃碼領(lǐng)資料
掃碼領(lǐng)資料掃碼領(lǐng)資料

電話:19522948401
-

 文獻(xiàn)獎勵申請
文獻(xiàn)獎勵申請