
蛋白-核酸互作研究方法
信息來源:金開瑞 作者:genecreate 發布時間:2017-11-30 16:05:33
蛋白質與RNA的相互作用是許多細胞功能的核心,如蛋白質合成、mRNA組裝、病毒復制、細胞發育調控等。蛋白-核酸互作是機制研究的重要組成部分,是表現、功能等研究的進一步深化。今天小編以一篇經典文獻帶你了解蛋白-核酸互作研究方法。
題目:Role of MYC-Regulated Long Noncoding RNAs in Cell Cycle Regulation and Tumorigenesis
期刊:Journal of the National Cancer Institute (JNCI)
影響因子:12.589
主要技術:RNA pull-down , LC-MS/MS, ChIP, RIP
研究背景
哺乳動物基因組編碼大量的非編碼RNA,如microRNAs、piRNAs和lncRNAs。迄今為止,已經注釋或鑒定了3萬多個lncRNAs。目前已經在幾種癌癥中鑒定到lncRNAs,在結直腸癌(CRC)中也有一些報道,如CCAT1和CCAT2,但大部分lncRNAs 在CRC中的作用仍然鮮為人知,有待闡明。已知轉錄因子MYC調節lncRNAs并且與癌細胞增殖和腫瘤發生相關。本文作者利用熒光素酶啟動子測定,ChIP,RNA pull-down,缺失圖譜測定,LC-MS / MS和RIP等方法手段來探索MYC調控lncRNAs的相關機制。
研究內容及結果
1. 鑒定結直腸癌(CRC)中特異表達的lncRNAs
作者利用lncRNAs基因芯片來分析正常結腸來源和CRC來源的細胞和組織中的lncRNAs。結果獲得30215個蛋白質編碼轉錄本和33045個有注釋或已知的lncRNAs。其中在CRC中具有表達差異的蛋白質編碼轉錄本(2212/30215)和lncRNAs(2325/33045)具有相似的比例,均為7%(圖1,A和B)。
在2325個差異表達的lncRNAs中,有1265個上調和1060下調(圖1C)。作者選取差異倍數大于15倍,P <0.01的差異表達的lncRNAs進行RACE快速擴增, 然后利用qRT-PCR進行驗證。結果有4個lncRNAs(CCAT3、CCAT4、CCAT5和CCAT6)在CRC細胞系中上調表達(圖1, D-G),CCAT7和CCAT8在CRC細胞系中下調表達(圖1,H-I)。利用52個CRC組織和它們臨近的正常組織(NATs)進行驗證也證實了上述結果(圖J-O)。
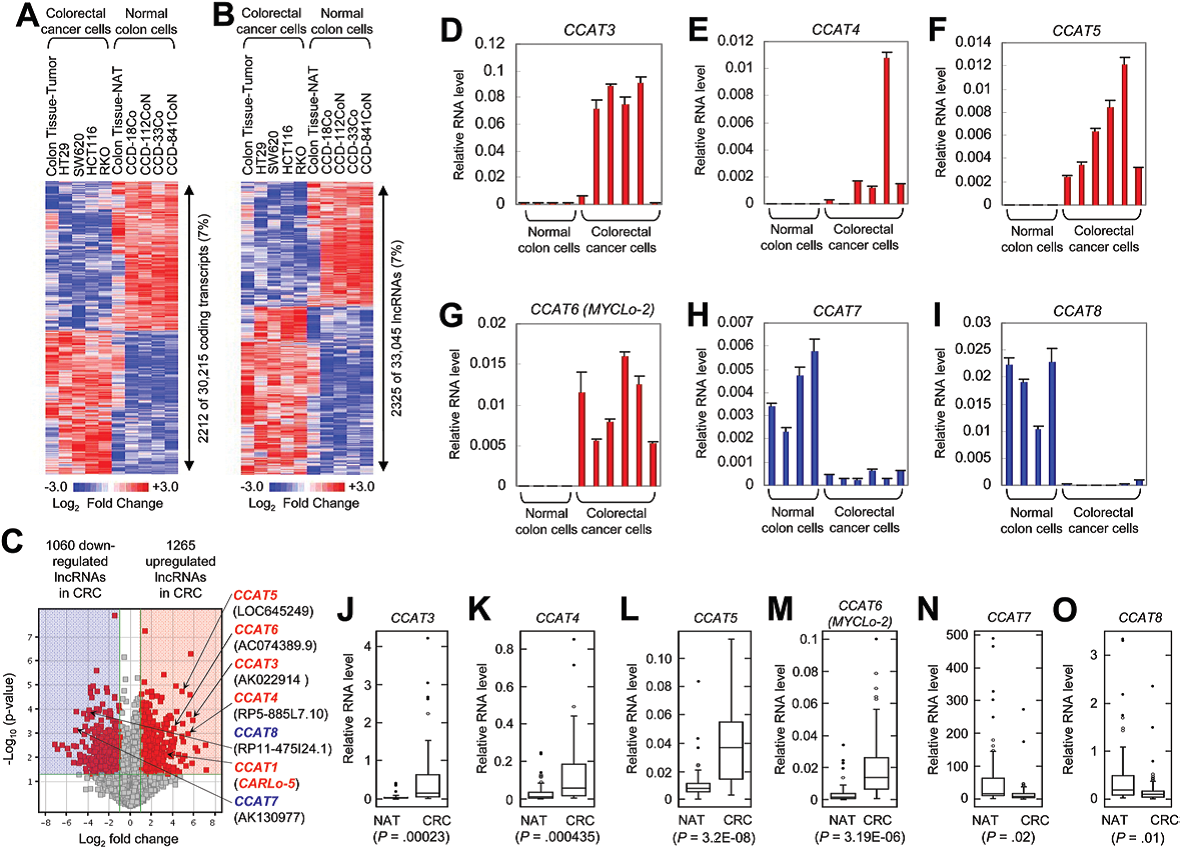
圖1 鑒定結腸直腸癌(CRC)中差異表達的lncRNAs
2. 進一步鑒定原癌基因MYC調控的CRC相關lncRNAs
為進一步鑒定由MYC調控的CRC相關lncRNAs的差異表達,作者利用lncRNAs微陣列篩選siMYC處理的CRC派生細胞系中的lncRNAs。結果,在HCT116細胞系和RKO細胞系中分別鑒定到324、863個由MYC調節的lncRNAs(圖2B,C)。通過進一步篩選獲得MYC上調的三種lncRNAs(AK021907,AC074389.9和KTN1-AS1)(圖2D)。這些由MYC調控的lncRNAs 被命名為MYCLos(MYCLos-1,MYCLos-2,MYCLos-3)。
接下來作者研究了由MYC介導調控的MYCLos在各種癌細胞中是否常見。結果表明,在各種癌癥類型的細胞中都能觀察到MYC介導調控MYCLos,表明MYC介導調控MYCLos在多種類型的癌癥中是保守的(圖2E-G)。
為了進一步驗證MYC與MYCLos之間的關系,作者還研究了50個人類原發性結直腸癌組織樣本(25個正常結腸和25個CRC組織樣本)的表達水平,根據MYC表達水平分為4個組(圖2H)。通過比較四組中MYCLo-1,-2和-3的表達水平,發現MYCLo-1,-2和-3的表達水平與MYC的表達水平在統計學上顯著相關(圖2,I-K)。此外,MYCLos在含有MYC過表達的CRC細胞系(圖1G)中高度表達(圖2A)。為測試MYC是否在轉錄水平調節MYCLo-1,-2和-3,作者利用熒光素酶報道分子對其進行了測定。結果表明,MYC基因抑制后,MYCLos啟動子的活性降低,表明MYC在轉錄水平上誘導MYCLos表達(圖2,L-N)。這些結果表明MYC誘導的MYCLos在轉錄水平上由轉錄增強子MYC直接調節。
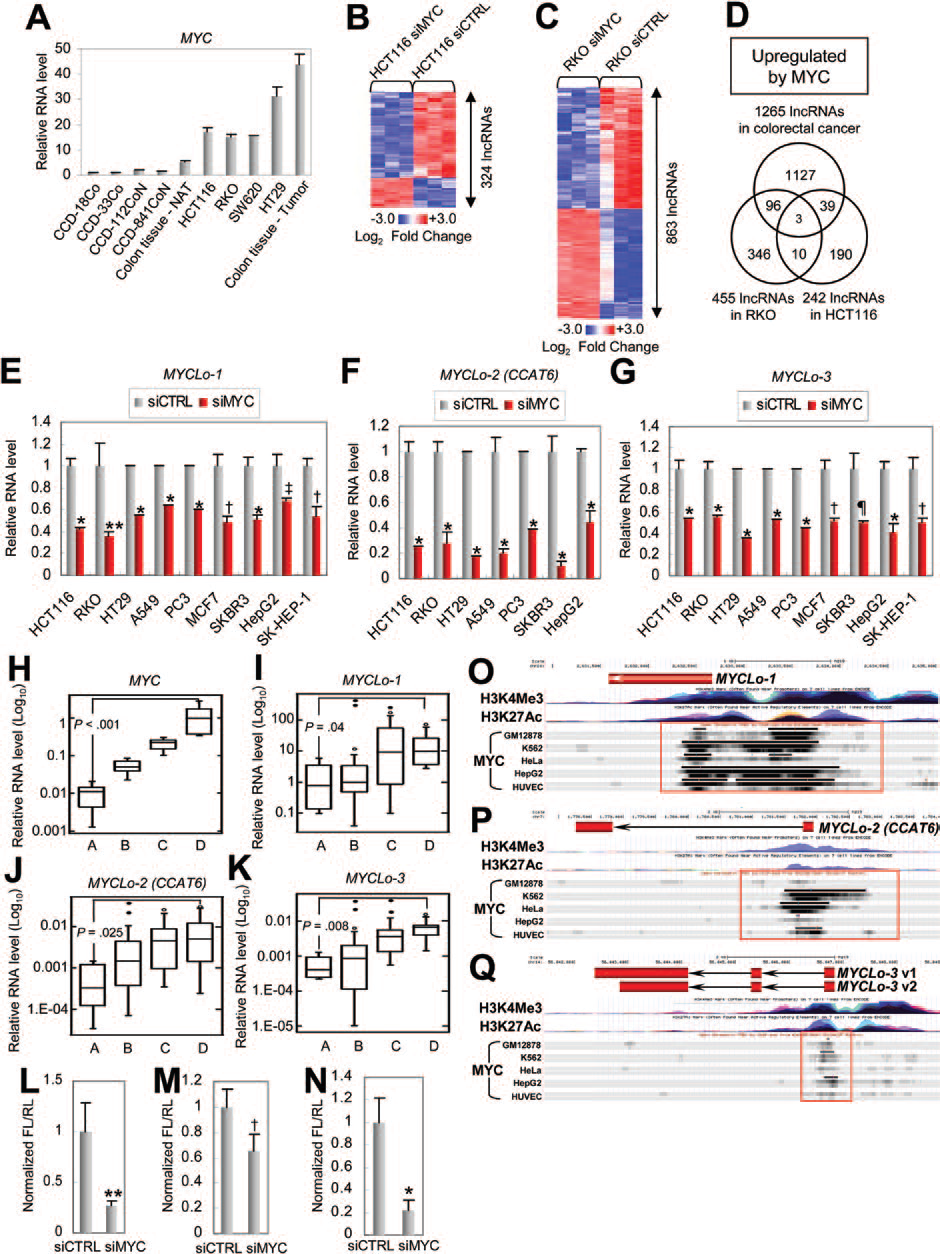
圖2 鑒定由MYC直接調控的CRC相關的lncRNAs
3. MYC誘導的MYCLos對CDKN1A、CDKN2B等MYC靶基因細胞增殖和表達的影響
原癌基因MYC通過調節許多細胞周期調節基因來促進細胞的增殖。作者將MYCLo-1,-2或-3以及MYC進行敲除,結果細胞增殖減少(圖3A,3B)。敲除MYCLo-1或-2導致細胞在G1期積累,表明MYCLo-1和-2參與G1 / S轉換(圖3D)。敲除MYCLo-3導致S和G2期細胞積累,表明MYCLo-3在G2期具有調節功能(圖3D)。另外作者還鑒定到有190、49和13個細胞周期調節基因分別由MYCLo-1、MYCLo-2和MYCLo-3調控(圖3E-F)。
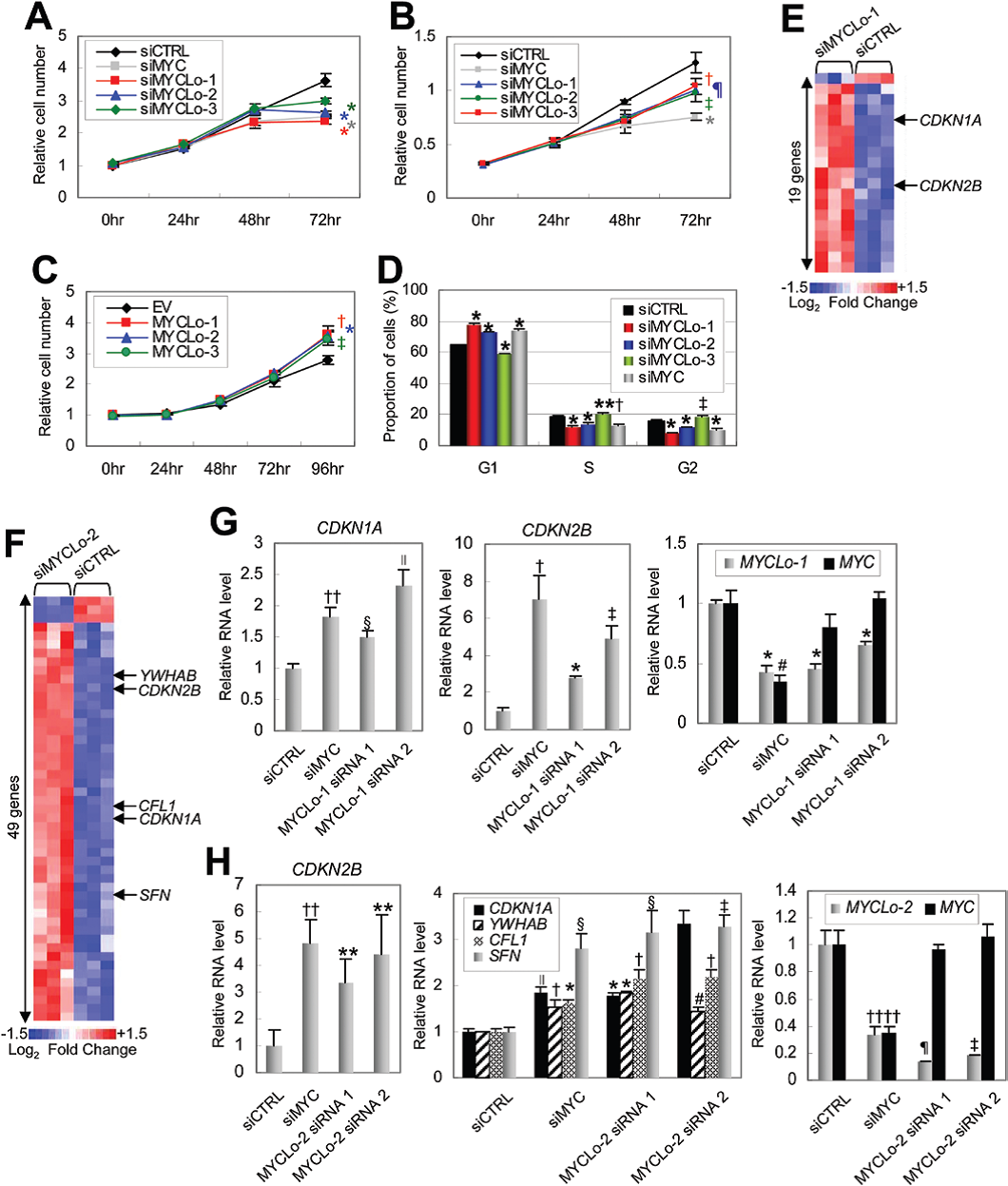
圖3 MYCLos敲除對細胞增殖和細胞周期進程的影響
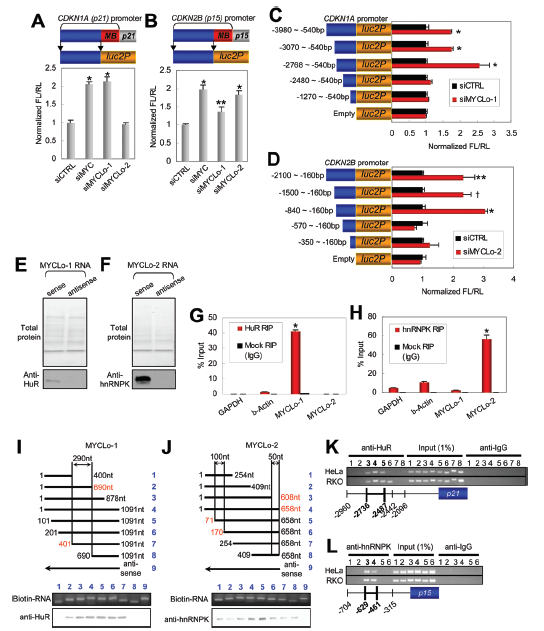
4. MYCLos介導的CDKN1A和CDKN2B在遠端啟動子區域的轉錄調節
為闡明MYC調控的lncRNAs抑制CDKN1A和CDKN2B表達的機制,作者分別在CDKN1A和CDKN2B基因的啟動子中擴增了兩個不同的區域,一個包括近端MYC結合(MB)區域,另一個包括沒有MB區域的上游區域。然后利用熒光素酶實驗對擴增的區域進行啟動子測試(圖4A和B)。MYC的敲除能誘導無MB區域的上游區域的啟動子活性,敲除MYCLo-1或 -2也能誘導激活CDKN1A 和 CDKN2B的啟動子活性(圖4A,B)。表明MYC不僅在近端區域而且在其啟動子的遠端區域也可以調控CDKN1A和CDKN2B的轉錄活性。進一步見鑒定結果表明CDKN1A啟動子區域大約在2768bp和2480bp之間對MYCLo-1調節CDKN1A起關鍵的作用(圖4C),CDKN2B啟動子區域大約在840bp和570bp之間對MYCLo-2調節CDKN2B起關鍵作用(圖4D)。
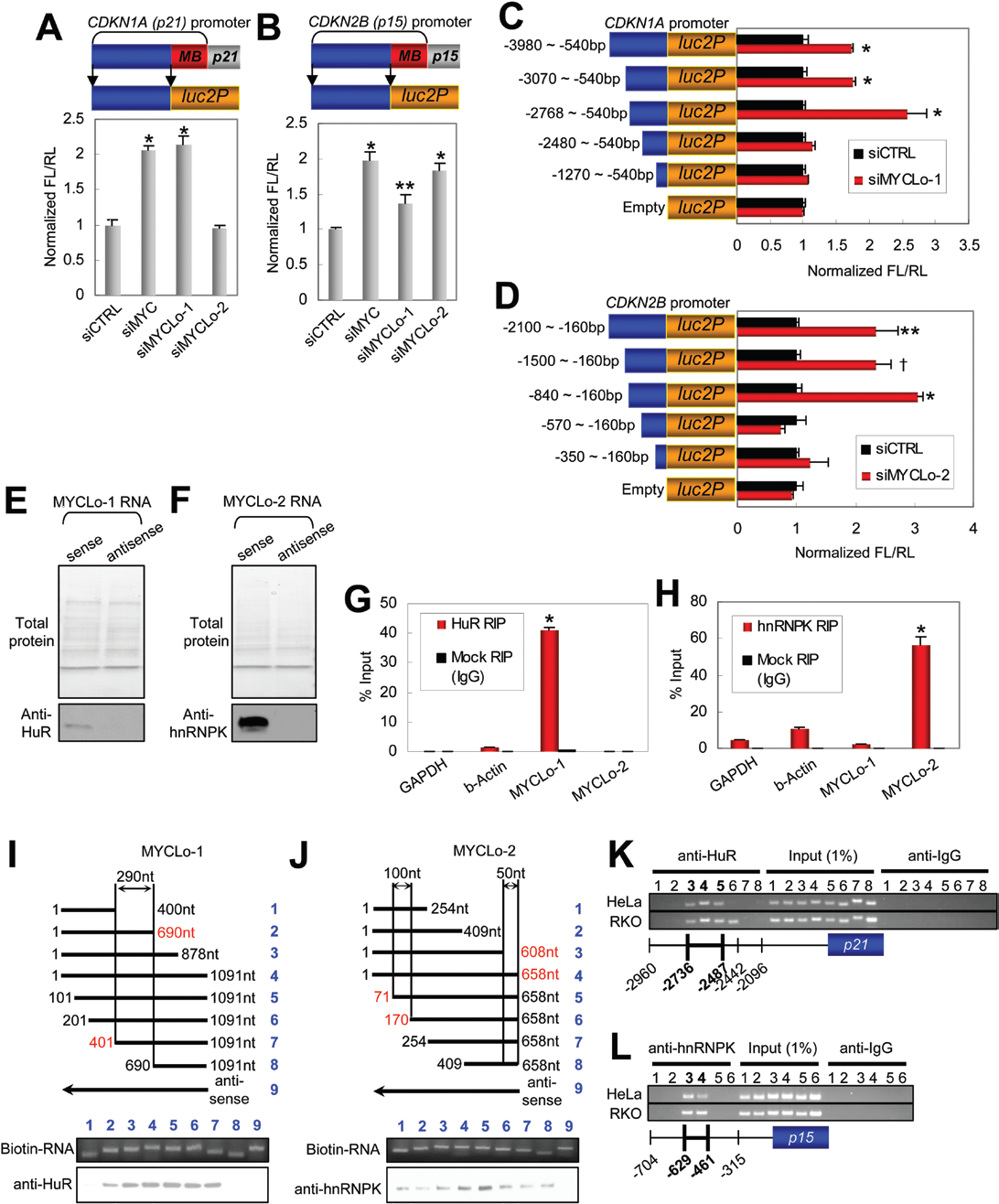
圖4 MYCLo-1/2和RNP結合蛋白之間的相互作用及其參與CDKN1A和CDKN2B的5'啟動子活性的調節
5. MYCLos和RNA結合蛋白(如HuR和hnRNPK)的相互作用
為研究MYCLos的功能機制,作者使用RNA pull-down實驗及質譜分析的方式鑒定RNA-相關的總蛋白(圖4E,F)。結果發現10個與MYCLo-1相關的蛋白和8個與MYCLo-2相關的蛋白。通過使用RPISeq軟件進行預測,作者還發現HuR-MYCLo-1、hnRNPK-MYCLo-2具有極大相互作用的可能性(圖4G,H)。并且通過Western blot驗證其相互作用(圖4E,F)。
為進一步確認HuR-MYCLo-1、hnRNPK-MYCLo-2之間的相互作用,作者利用RIP實驗進行了驗證。結果與RNA pull-down實驗結果一致,使用HuR抗體的RIP樣品中MYCLo-1獲得了富集(圖4G),使用hnRNPK抗體的RIP樣品中MYCLo-2也獲得了富集(圖4H)。這些結果表明HuR與MYCLo1的交互作用與CDKN1A的轉錄抑制有關,MYCLo-2通過與hnRNPK相互作用抑制CDKN2B的轉錄。
6. MYCLo-2的致癌功能
作者利用Northern blot驗證了CRC細胞系中MYCLo-2(CCAT6)的過表達。發現大多數CRC組織中MYCLo-2具有非常高的表達水平,在52組相比較的組織樣品中有46個CRC組織的MYCLo-2高度表達(圖5A)。此外,與正常前列腺來源的細胞相比,MYCLo-2也在前列腺癌(PC)細胞和組織中過表達(圖5B, C)。在體外集落形成測定中,敲除MYCLo-2能顯著減少細胞和集落的數量,表明MYCLo-2在腫瘤細胞轉化中具有作用(圖5D)。
另外作者還通過使用MYCLo-2 siRNAs轉染腫瘤細胞的體內異種移植,研究了MYCLo-2在腫瘤發生中的抑制作用。結果表明,MYCLo-2損耗對腫瘤的發展有明顯的抑制作用(圖5E)。上述結果表明,MYCLo-2不僅在腫瘤中過表達,還參與腫瘤發生和腫瘤生長。
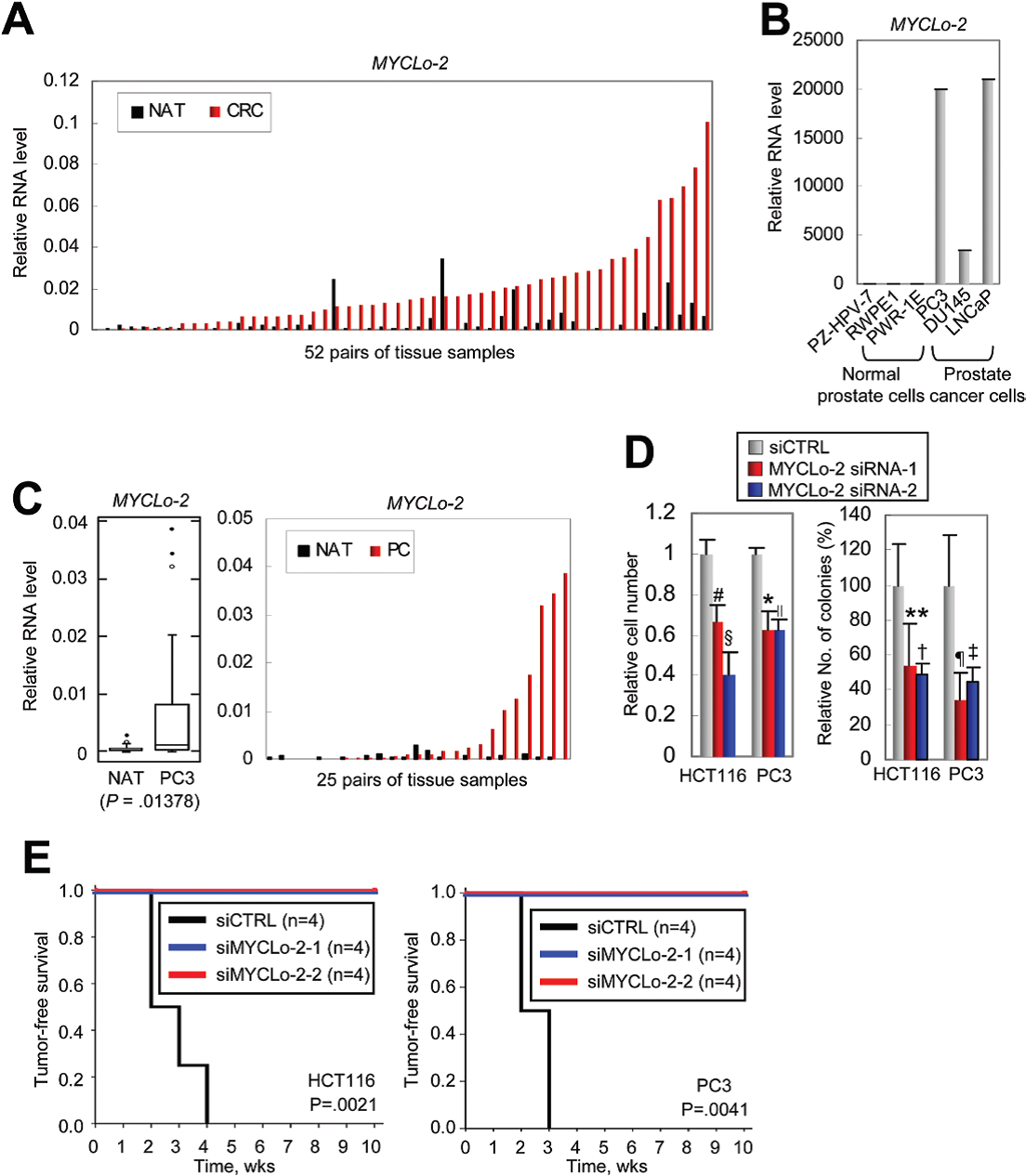
圖5 MYCLo-2在癌細胞轉化和腫瘤發生中的功能
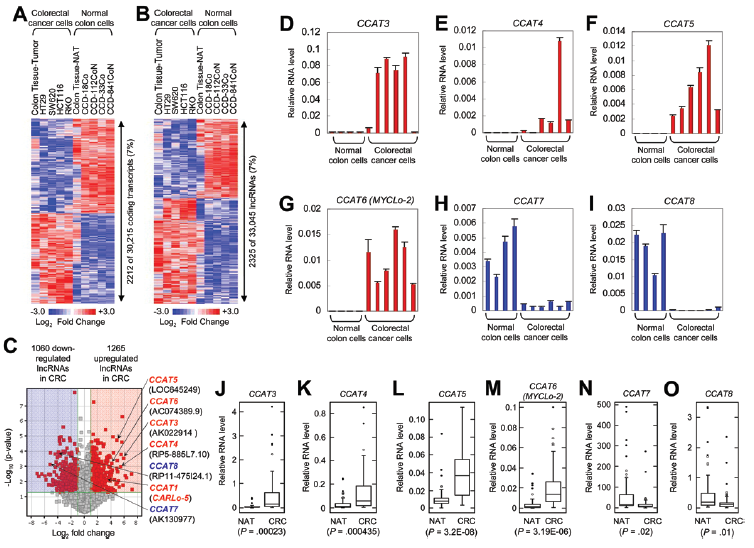
圖1 鑒定結腸直腸癌(CRC)中差異表達的lncRNAs
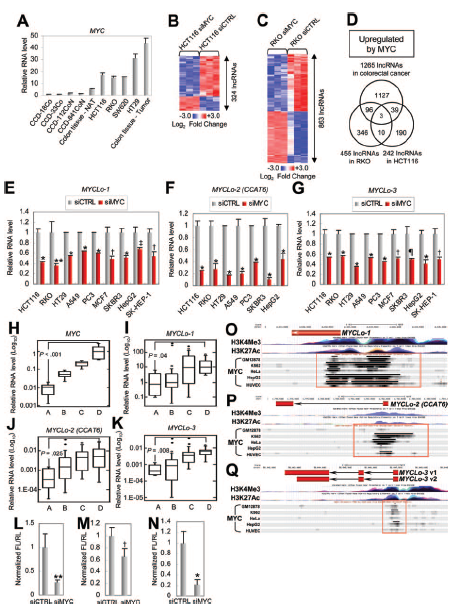
圖2 鑒定由MYC直接調控的CRC相關的lncRNAs
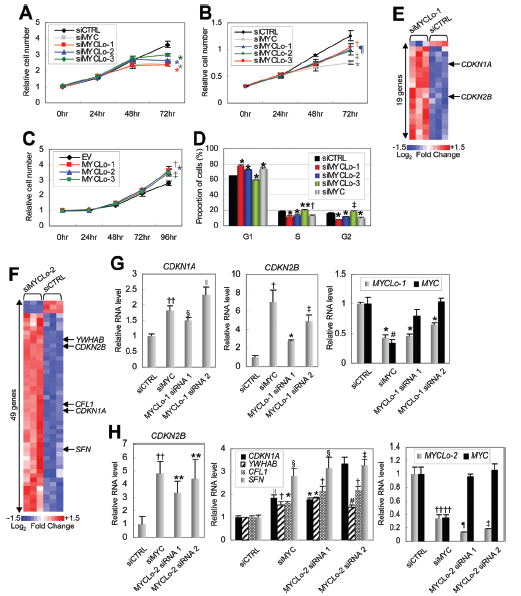
圖3 MYCLos敲除對細胞增殖和細胞周期進程的影響
經典文獻解讀:MYC調控的長鏈非編碼RNA(lncRNAs)在細胞周期調控和腫瘤發生中的作用
基本信息題目:Role of MYC-Regulated Long Noncoding RNAs in Cell Cycle Regulation and Tumorigenesis
期刊:Journal of the National Cancer Institute (JNCI)
影響因子:12.589
主要技術:RNA pull-down , LC-MS/MS, ChIP, RIP
研究背景
哺乳動物基因組編碼大量的非編碼RNA,如microRNAs、piRNAs和lncRNAs。迄今為止,已經注釋或鑒定了3萬多個lncRNAs。目前已經在幾種癌癥中鑒定到lncRNAs,在結直腸癌(CRC)中也有一些報道,如CCAT1和CCAT2,但大部分lncRNAs 在CRC中的作用仍然鮮為人知,有待闡明。已知轉錄因子MYC調節lncRNAs并且與癌細胞增殖和腫瘤發生相關。本文作者利用熒光素酶啟動子測定,ChIP,RNA pull-down,缺失圖譜測定,LC-MS / MS和RIP等方法手段來探索MYC調控lncRNAs的相關機制。
研究內容及結果
1. 鑒定結直腸癌(CRC)中特異表達的lncRNAs
作者利用lncRNAs基因芯片來分析正常結腸來源和CRC來源的細胞和組織中的lncRNAs。結果獲得30215個蛋白質編碼轉錄本和33045個有注釋或已知的lncRNAs。其中在CRC中具有表達差異的蛋白質編碼轉錄本(2212/30215)和lncRNAs(2325/33045)具有相似的比例,均為7%(圖1,A和B)。
在2325個差異表達的lncRNAs中,有1265個上調和1060下調(圖1C)。作者選取差異倍數大于15倍,P <0.01的差異表達的lncRNAs進行RACE快速擴增, 然后利用qRT-PCR進行驗證。結果有4個lncRNAs(CCAT3、CCAT4、CCAT5和CCAT6)在CRC細胞系中上調表達(圖1, D-G),CCAT7和CCAT8在CRC細胞系中下調表達(圖1,H-I)。利用52個CRC組織和它們臨近的正常組織(NATs)進行驗證也證實了上述結果(圖J-O)。
圖1 鑒定結腸直腸癌(CRC)中差異表達的lncRNAs
2. 進一步鑒定原癌基因MYC調控的CRC相關lncRNAs
為進一步鑒定由MYC調控的CRC相關lncRNAs的差異表達,作者利用lncRNAs微陣列篩選siMYC處理的CRC派生細胞系中的lncRNAs。結果,在HCT116細胞系和RKO細胞系中分別鑒定到324、863個由MYC調節的lncRNAs(圖2B,C)。通過進一步篩選獲得MYC上調的三種lncRNAs(AK021907,AC074389.9和KTN1-AS1)(圖2D)。這些由MYC調控的lncRNAs 被命名為MYCLos(MYCLos-1,MYCLos-2,MYCLos-3)。
接下來作者研究了由MYC介導調控的MYCLos在各種癌細胞中是否常見。結果表明,在各種癌癥類型的細胞中都能觀察到MYC介導調控MYCLos,表明MYC介導調控MYCLos在多種類型的癌癥中是保守的(圖2E-G)。
為了進一步驗證MYC與MYCLos之間的關系,作者還研究了50個人類原發性結直腸癌組織樣本(25個正常結腸和25個CRC組織樣本)的表達水平,根據MYC表達水平分為4個組(圖2H)。通過比較四組中MYCLo-1,-2和-3的表達水平,發現MYCLo-1,-2和-3的表達水平與MYC的表達水平在統計學上顯著相關(圖2,I-K)。此外,MYCLos在含有MYC過表達的CRC細胞系(圖1G)中高度表達(圖2A)。為測試MYC是否在轉錄水平調節MYCLo-1,-2和-3,作者利用熒光素酶報道分子對其進行了測定。結果表明,MYC基因抑制后,MYCLos啟動子的活性降低,表明MYC在轉錄水平上誘導MYCLos表達(圖2,L-N)。這些結果表明MYC誘導的MYCLos在轉錄水平上由轉錄增強子MYC直接調節。
圖2 鑒定由MYC直接調控的CRC相關的lncRNAs
原癌基因MYC通過調節許多細胞周期調節基因來促進細胞的增殖。作者將MYCLo-1,-2或-3以及MYC進行敲除,結果細胞增殖減少(圖3A,3B)。敲除MYCLo-1或-2導致細胞在G1期積累,表明MYCLo-1和-2參與G1 / S轉換(圖3D)。敲除MYCLo-3導致S和G2期細胞積累,表明MYCLo-3在G2期具有調節功能(圖3D)。另外作者還鑒定到有190、49和13個細胞周期調節基因分別由MYCLo-1、MYCLo-2和MYCLo-3調控(圖3E-F)。
圖3 MYCLos敲除對細胞增殖和細胞周期進程的影響
4. MYCLos介導的CDKN1A和CDKN2B在遠端啟動子區域的轉錄調節
為闡明MYC調控的lncRNAs抑制CDKN1A和CDKN2B表達的機制,作者分別在CDKN1A和CDKN2B基因的啟動子中擴增了兩個不同的區域,一個包括近端MYC結合(MB)區域,另一個包括沒有MB區域的上游區域。然后利用熒光素酶實驗對擴增的區域進行啟動子測試(圖4A和B)。MYC的敲除能誘導無MB區域的上游區域的啟動子活性,敲除MYCLo-1或 -2也能誘導激活CDKN1A 和 CDKN2B的啟動子活性(圖4A,B)。表明MYC不僅在近端區域而且在其啟動子的遠端區域也可以調控CDKN1A和CDKN2B的轉錄活性。進一步見鑒定結果表明CDKN1A啟動子區域大約在2768bp和2480bp之間對MYCLo-1調節CDKN1A起關鍵的作用(圖4C),CDKN2B啟動子區域大約在840bp和570bp之間對MYCLo-2調節CDKN2B起關鍵作用(圖4D)。
圖4 MYCLo-1/2和RNP結合蛋白之間的相互作用及其參與CDKN1A和CDKN2B的5'啟動子活性的調節
5. MYCLos和RNA結合蛋白(如HuR和hnRNPK)的相互作用
為研究MYCLos的功能機制,作者使用RNA pull-down實驗及質譜分析的方式鑒定RNA-相關的總蛋白(圖4E,F)。結果發現10個與MYCLo-1相關的蛋白和8個與MYCLo-2相關的蛋白。通過使用RPISeq軟件進行預測,作者還發現HuR-MYCLo-1、hnRNPK-MYCLo-2具有極大相互作用的可能性(圖4G,H)。并且通過Western blot驗證其相互作用(圖4E,F)。
為進一步確認HuR-MYCLo-1、hnRNPK-MYCLo-2之間的相互作用,作者利用RIP實驗進行了驗證。結果與RNA pull-down實驗結果一致,使用HuR抗體的RIP樣品中MYCLo-1獲得了富集(圖4G),使用hnRNPK抗體的RIP樣品中MYCLo-2也獲得了富集(圖4H)。這些結果表明HuR與MYCLo1的交互作用與CDKN1A的轉錄抑制有關,MYCLo-2通過與hnRNPK相互作用抑制CDKN2B的轉錄。
6. MYCLo-2的致癌功能
作者利用Northern blot驗證了CRC細胞系中MYCLo-2(CCAT6)的過表達。發現大多數CRC組織中MYCLo-2具有非常高的表達水平,在52組相比較的組織樣品中有46個CRC組織的MYCLo-2高度表達(圖5A)。此外,與正常前列腺來源的細胞相比,MYCLo-2也在前列腺癌(PC)細胞和組織中過表達(圖5B, C)。在體外集落形成測定中,敲除MYCLo-2能顯著減少細胞和集落的數量,表明MYCLo-2在腫瘤細胞轉化中具有作用(圖5D)。
另外作者還通過使用MYCLo-2 siRNAs轉染腫瘤細胞的體內異種移植,研究了MYCLo-2在腫瘤發生中的抑制作用。結果表明,MYCLo-2損耗對腫瘤的發展有明顯的抑制作用(圖5E)。上述結果表明,MYCLo-2不僅在腫瘤中過表達,還參與腫瘤發生和腫瘤生長。
圖5 MYCLo-2在癌細胞轉化和腫瘤發生中的功能
文章小結
該文章使用多種技術研究MYC調控lncRNAs的相關機制,揭示了MYCLos通過調節已知的MYC靶基因的表達來參與細胞增殖和細胞周期的調控,另外還發現MYCLo-1與HuR和MYCLo-2與hnRNPK之間的相互作用。進一步研究表明這些相互作用可能參與對MYC靶標(如CDKN1A和CDKN2B)的調控。此外,將MYCLo-2進行敲除能夠抑制癌細胞轉化和腫瘤發生。這些研究成果對于發現新的癌癥靶標和研究MYC相關的發病機制具有重要意義。文獻來源
Kim, T., Jeon, Y. J., Cui, R., Lee, J. H., Peng, Y., & Kim, S. H., et al. (2015). Role of myc-regulated long noncoding rnas in cell cycle regulation and tumorigenesis. Journal of the National Cancer Institute, 107(4), dju505.
圖1 鑒定結腸直腸癌(CRC)中差異表達的lncRNAs
圖2 鑒定由MYC直接調控的CRC相關的lncRNAs
圖3 MYCLos敲除對細胞增殖和細胞周期進程的影響
圖4 MYCLo-1/2和RNP結合蛋白之間的相互作用及其參與CDKN1A和CDKN2B的5'啟動子活性的調節
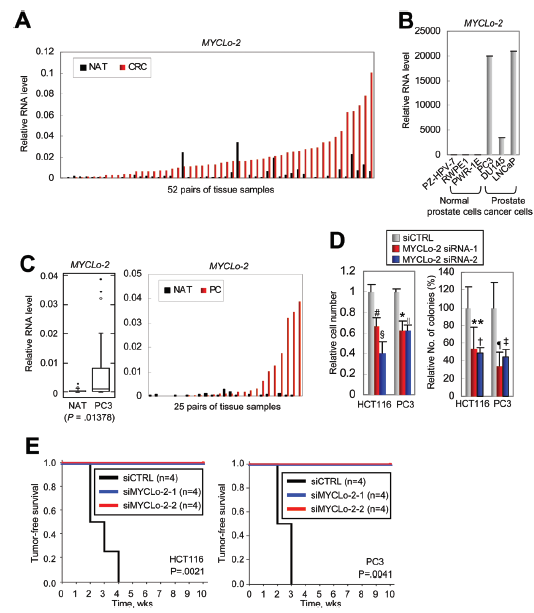
圖5 MYCLo-2在癌細胞轉化和腫瘤發生中的功能
圖5 MYCLo-2在癌細胞轉化和腫瘤發生中的功能
上一條:腺病毒包裝簡介
下一條:金開瑞帶您走近免疫共沉淀
最新動態
-
09.23
中藥的現代詮釋:外泌體如何革新傳統醫學?
-
07.02
1+1>2!深度解析RNA測序數據挖掘邏輯和后期實驗設計思路,輕松研獲10+ SCI
-
07.01
“稻”亦有道——盤點近期水稻研究的重大突破
-
06.28
科學與美學的結合體:植物亞細胞定位技術詳解
-
06.28
“聚焦新質生產力,激發科研新動能”|LCA躋身蛋白互作研究的新銳力量
-
06.05
知無不“研”|一文讀懂免疫共沉淀技術(Co-IP)
-
05.14
四大研究利器(Co-IP、BIFC、Y2H、GST pull-down)助力速配蛋白互作“最佳拍檔”
-
05.14
高效、精準、直觀、實時——取經“蛋白互作研究翹楚”BIFC!
-
05.14
轉染效率低、干擾效果差、重復性欠佳...siRNA研究頻遇“攔路虎”怎么辦?
-
04.22
一文讀懂EMSA技術核心要點,讓“emsa” 秒變“easy”
 X
X 






