
Nature 揭示真核生物細(xì)胞核中染色質(zhì)分離新機(jī)制:異染色質(zhì)驅(qū)動(dòng)倒置核和常規(guī)核的分隔
信息來源:金開瑞 作者:genecreate 發(fā)布時(shí)間:2019-07-18 16:18:11
題目:Heterochromatin drives compartmentalization of inverted and conventional nuclei
期刊:Nature
影響因子:41.577
主要技術(shù):Hi-C、Cryosections 、immunostaining、FISH 、microscopy
哺乳動(dòng)物細(xì)胞中常染色質(zhì)和異染色質(zhì)存在空間上的分離,但區(qū)室化的形成機(jī)制尚不清楚。本研究對(duì)夜行哺乳動(dòng)物的視桿細(xì)胞倒置核進(jìn)行Hi-C分析并結(jié)合顯微鏡和多聚體模擬,發(fā)現(xiàn)異染色質(zhì)區(qū)域間的吸引對(duì)建立區(qū)室化及著絲粒周邊異染色質(zhì),可調(diào)節(jié)性異染色質(zhì)和常染色質(zhì)的建立至關(guān)重要。倒置細(xì)胞核中加入異染色質(zhì)和核纖層的相互作用可重建傳統(tǒng)細(xì)胞核中的組織。
文章思路
研究?jī)?nèi)容及結(jié)果
1. 常規(guī)和倒置核的顯微鏡觀察及Hi-C分析
為了研究基因組區(qū)室化的機(jī)制,作者對(duì)初生組織中分離的四種小鼠細(xì)胞類型中進(jìn)行了Hi-C實(shí)驗(yàn)。這四種細(xì)胞類型均具有常規(guī)或倒置核結(jié)構(gòu):桿光感受器(倒置),非桿視神經(jīng)元細(xì)胞(常規(guī)),野生型胸腺細(xì)胞(常規(guī))和無效核纖層蛋白B受體胸腺細(xì)胞(Lbr - / -)(倒置)(Fig. 1a)。常規(guī)核和倒置核從顯微鏡中可以看出核組織的巨大差異(Fig.1a),但是染色質(zhì)組織特征中的拓?fù)浣Y(jié)合域(TAD),染色體區(qū)域和區(qū)室均類似(Fig1.b),在單細(xì)胞Hi-C中也發(fā)現(xiàn)上述特點(diǎn)。
隨后,作者研究了常染色質(zhì)和異染色質(zhì)在空間定位上的差異是否會(huì)影響Hi-C中所見的核區(qū)隔,從Hi-C圖中計(jì)算了隔室剖面(Fig.1b),并將隔室化程度定義為隔室之間接觸的富集程度。在倒置核中,胸腺細(xì)胞的分隔程度僅略有降低,但在桿狀細(xì)胞中則變得更強(qiáng)(Fig.1c)。
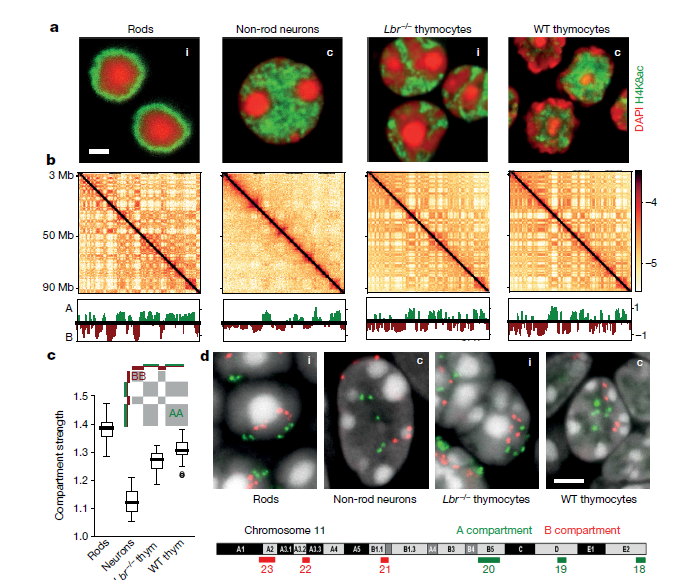
Fig.1常規(guī)和倒置核的顯微鏡觀察和Hi-C分析
綜上所述,分析表明,盡管在倒置時(shí)單個(gè)A或B室的空間位置發(fā)生了變化,但仍保留了分區(qū)的程度(Fig.1a, d),表明其分區(qū)的機(jī)制不能嚴(yán)格依賴于核層。為了協(xié)調(diào)倒置核與常規(guī)核的Hi-c區(qū)室的劃分與這些核中不同空間幾何形狀之間的關(guān)系,作者尋求一種滿足以下三個(gè)標(biāo)準(zhǔn)的劃分機(jī)制。1、它應(yīng)該重現(xiàn)倒置的核,通過顯微鏡定量定義不同類型染色質(zhì)的徑向位置和Hi-C技術(shù)區(qū)分區(qū)室化的強(qiáng)度。2、它應(yīng)該重現(xiàn)常規(guī)的核,當(dāng)引入異染色質(zhì)和核層之間的有吸引力的相互作用時(shí),常規(guī)的核的特征是在Hi-C中顯示類似程度的區(qū)室化,但是在顯微鏡觀察下區(qū)室的空間位置明顯不同。3、它應(yīng)該是基于合理的生物上和物理上的力,這就限制了不同類型的染色質(zhì)與核層的染色質(zhì)之間的相互作用。
2. 倒置的核的形態(tài)限制了可能的區(qū)室化模型
為測(cè)試區(qū)室化的機(jī)制,作者開發(fā)了染色質(zhì)的平衡聚合物模型,其代表染色體作為嵌段共聚物(Fig.2a),類似于其他分離的分離模型。擴(kuò)展以前的兩種模型,作者模擬使用三種類型的單體:常染色質(zhì)(A)異染色(B)和中心體組成型異染色質(zhì)(C)。作者模擬了8條染色體;每條染色體由6,000個(gè)單體組成;每個(gè)單體代表40kb染色質(zhì);在球形核中以35%的體積密度分布。A和B單體的序列反映倒置的核的Hi-C數(shù)據(jù)中的區(qū)室化。為了表示Hi-C中無法顯示的中心區(qū)域或染色體中心,作者在每個(gè)染色體近端放置了1塊C單體(長(zhǎng)度為染色體的16%)。所有單體都排除了體積,并根據(jù)其染色質(zhì)類型的短距離成對(duì)吸引。給定六個(gè)成對(duì)吸引參數(shù)(A-A,A-B,B-B,B-C,C-C和A-C),所有可能的吸引強(qiáng)度排列都指定720(6)類模型(參見方法)。為了限制可能模型的空間,作者首先定量地將所有720類模型與顯微鏡觀察的數(shù)據(jù)進(jìn)行比較。 具體而言,作者計(jì)算了A,B和C單體的徑向分布,并將模擬中獲得的分布與顯微鏡中獲得的分布進(jìn)行了比較(Fig.2b)。大多數(shù)模型類型與顯微鏡中觀察到的倒置核的同心幾何形狀不一致(Fig.2c)。 例如,過強(qiáng)的B-C相互作用導(dǎo)致B和C混合(Fig.2c,模型8和擴(kuò)展數(shù)據(jù)Fig.6a-c),而相對(duì)較弱的B-C相互作用導(dǎo)致C單體染色體從a中排出B單體的中心質(zhì)量(Fig.2c,模型112)。過強(qiáng)的A-A相互作用傾向于促進(jìn)大的常染色質(zhì)小球的形成(Fig.2c,模型650和擴(kuò)展數(shù)據(jù)Fig.6d-f)。
以上結(jié)果表明,與常染色區(qū)域的活動(dòng)相關(guān)的聚類是作為區(qū)域化基礎(chǔ)的主要機(jī)制。只有八類模型可以重現(xiàn)實(shí)驗(yàn)觀察到的倒置幾何形狀(Fig.2b,c)。作者專注于最合適的模型類型,并進(jìn)一步簡(jiǎn)化這些模型,將C-C固定得足夠高以誘導(dǎo)C單體的中心球,A-A始終是遠(yuǎn)小于B-B(擴(kuò)展數(shù)據(jù)Fig.7d),并且所有交叉項(xiàng)都是各個(gè)純項(xiàng)的幾何平均值(例如,A-B =(A-A×B-B)1/2) ,從而滿足Flory-Huggins相分離標(biāo)準(zhǔn)。 這使得B-B吸引力成為唯一的自由參數(shù)。
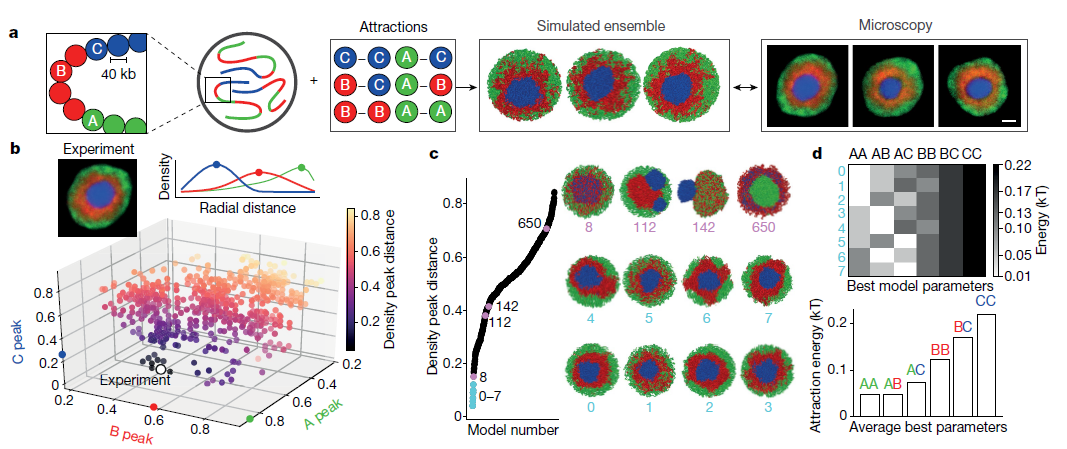
Fig.2 倒置的核的形態(tài)限制了可能的區(qū)室化模型
3. 基于異染色質(zhì)的機(jī)制定量地再現(xiàn)倒置和常規(guī)核
接下來,作者再現(xiàn)顯微鏡圖像中看到的反轉(zhuǎn)核的異染色質(zhì)主導(dǎo)模型,測(cè)試其是否可以同時(shí)再現(xiàn)在Hi-C數(shù)據(jù)中觀察到的區(qū)室化。通過確定相互作用強(qiáng)度的順序,作者發(fā)現(xiàn)了一系列B-B吸引的模型可以定量再現(xiàn)Hi-C數(shù)據(jù)和顯微鏡觀察(Fig.3a,b)。作者通過對(duì)倒置核的分析揭示的異染色區(qū)域之間的吸引力的中心作用,與此形成對(duì)比觀點(diǎn)認(rèn)為異染色區(qū)域間相互作用取決于同染色區(qū)域之間相互作用,或者是同作為分隔作用主要驅(qū)動(dòng)力是薄層之間相互作用的。異染色區(qū)域之間更強(qiáng)的吸引力與最近觀察到的異染色區(qū)域相關(guān)組蛋白甲基化在決定染色體力學(xué)性能方面的主導(dǎo)作用是一致的。
為了將作者的模型擴(kuò)展到傳統(tǒng)的原子核中,其表示了具有短程吸引的異染色質(zhì)-薄層相互作用(B-Lam引力,F(xiàn)ig.3c, d)。為了對(duì)實(shí)驗(yàn)中發(fā)現(xiàn)的不同染色體進(jìn)行建模,作者將c單體簇固定在薄層上隨機(jī)的位置。固定不需要在一段時(shí)間內(nèi)保持不同的染色體中心,但需要在平衡模擬中保持它們的分離(補(bǔ)充視頻1)。通過全面的B-B和B-Lam吸引物,作者發(fā)現(xiàn)他的模型可以同時(shí)重現(xiàn)顯微鏡圖像中觀察到的活性和非活性染色質(zhì)的空間定位,以及野生型胸腺細(xì)胞在Hi-C數(shù)據(jù)中觀察到的區(qū)室化。盡管再現(xiàn)顯微鏡數(shù)據(jù)需要足夠強(qiáng)的B-Lam而不進(jìn)一步限制這些參數(shù),同時(shí)再現(xiàn)在Hi-C數(shù)據(jù)中觀察到的區(qū)室化會(huì)縮小B-Lam和B-B吸引力的范圍(Fig.3c,d)。值得注意的是,常規(guī)核的最佳擬合B-B吸引區(qū)域包含最適合倒置核的B-B吸引力。由于組蛋白修飾與倒置和常規(guī)細(xì)胞核中相同類型的染色質(zhì)相關(guān),作者謹(jǐn)慎地假設(shè)B-B吸引力在兩種核類型中保持相同。通過這種約束,作者可以縮小可能的B-Lam值的范圍(約0.3kT;Fig.3c),并發(fā)現(xiàn)B-Lam吸引力應(yīng)該與B-B吸引力相當(dāng)。總之,作者的模擬表明兩者都是由倒置和常規(guī)細(xì)胞核主要由異染色質(zhì) - 異染色質(zhì)吸引力控制,而異染色質(zhì) - 薄層吸引力控制全局空間形態(tài)。
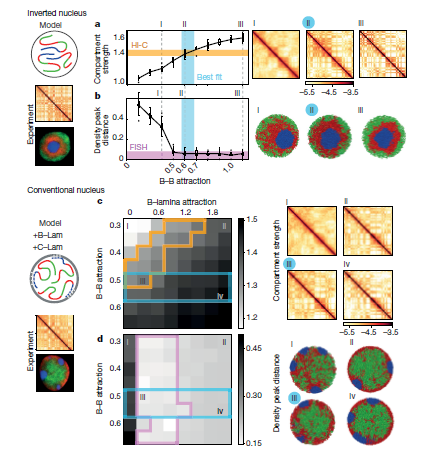
Fig.3異染色質(zhì)為基礎(chǔ)的機(jī)制定量復(fù)制倒置核和常規(guī)核
4. 隔室強(qiáng)度在核反演過程模型和實(shí)驗(yàn)中的時(shí)間演化和維持過程。
為了驗(yàn)證作者提出的劃分機(jī)制,作者模擬了一個(gè)核反轉(zhuǎn)的時(shí)間過程(Fig.4a, b),為此,作者關(guān)閉了模擬常規(guī)核中薄片-異染色質(zhì)的相互作用,并觀察到了自發(fā)核反轉(zhuǎn)(Fig.4b)。值得注意的是,模擬的時(shí)間過程反映了活躍的桿分化過程中的關(guān)鍵 (Fig.4b)。B和C單體在模擬中經(jīng)歷不可逆的液體狀融合,類似于其他相分離系統(tǒng)(Fig.4b)。在模擬中,雖然在異染色質(zhì)離開薄層后,區(qū)隔分離會(huì)暫時(shí)下降(Fig.4a),但在整個(gè)反轉(zhuǎn)過程中,區(qū)隔仍然是分離的。在顯微鏡顯示在棒狀的核轉(zhuǎn)化的整個(gè)過程期間,個(gè)體基因組位點(diǎn)與其自身區(qū)室類型的染色質(zhì)一起重新定位。例如,視紫紅質(zhì)基因座仍與常染色質(zhì)(A區(qū)室)相關(guān),并且視紫紅質(zhì)受體在整個(gè)翻轉(zhuǎn)過程中保持表達(dá)(Fig.4c)。為了進(jìn)一步測(cè)試作者提出的分區(qū)機(jī)制,從倒置幾何中初始化模擬并重新引入薄層-異染色質(zhì)相互作用。這些模擬預(yù)測(cè)只有部分去反轉(zhuǎn):而B單體取代了核周邊的A單體,C單體仍然是由B單體包圍并與薄層相關(guān)的單個(gè)大小球。
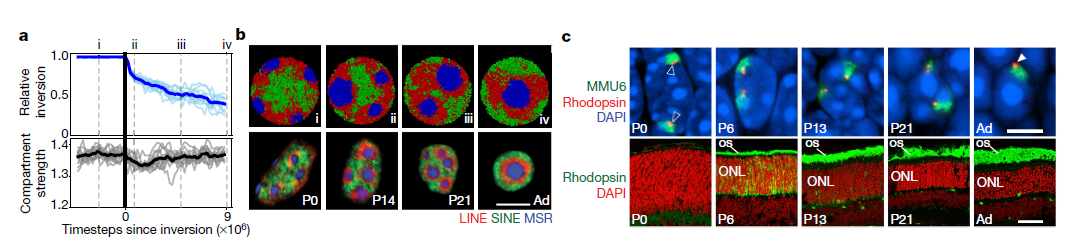
Fig.4 隔室強(qiáng)度在核反演過程模型和實(shí)驗(yàn)中的時(shí)間演化和維持過程
文章小結(jié)
哺乳動(dòng)物細(xì)胞中常染色質(zhì)和異染色質(zhì)存在空間上的分離,但區(qū)室化的形成機(jī)制尚不清楚。本研究對(duì)夜行哺乳動(dòng)物的視桿細(xì)胞倒置核進(jìn)行Hi-C分析并結(jié)合顯微鏡和多聚體模擬,發(fā)現(xiàn)異染色質(zhì)區(qū)域間的吸引對(duì)建立區(qū)室化及著絲粒周邊異染色質(zhì),可調(diào)節(jié)性異染色質(zhì)和常染色質(zhì)的建立至關(guān)重要。倒置細(xì)胞核中加入異染色質(zhì)和核纖層的相互作用可重建傳統(tǒng)細(xì)胞核中的組織。
解析文獻(xiàn)
Martin Falk, et al. Heterochromatin drives compartmentalization of inverted and conventional nuclei [J]. Nature, 2019, 570, pages395–399.
相關(guān)服務(wù)
Hi-C是以整個(gè)細(xì)胞核為研究對(duì)象,利用高通量測(cè)序技術(shù),結(jié)合生物信息分析方法,研究全基因組范圍內(nèi)整個(gè)染色質(zhì)DNA在空間位置上的關(guān)系,獲得高分辨率的染色質(zhì)調(diào)控元件相互作用圖譜。Hi-C可以與RNA-Seq、ChIP-Seq、ATAC-Seq等數(shù)據(jù)進(jìn)行聯(lián)合分析,從基因調(diào)控網(wǎng)絡(luò)和表觀遺傳網(wǎng)絡(luò)來闡述生物體性狀形成的相關(guān)機(jī)制。
武漢金開瑞生物工程有限公司提供的Hi-C技術(shù),是一種新型、高效、經(jīng)濟(jì)的研究方法,幫助您解析基因組三維結(jié)構(gòu)。
最新動(dòng)態(tài)
-
09.23
中藥的現(xiàn)代詮釋:外泌體如何革新傳統(tǒng)醫(yī)學(xué)?
-
07.02
1+1>2!深度解析RNA測(cè)序數(shù)據(jù)挖掘邏輯和后期實(shí)驗(yàn)設(shè)計(jì)思路,輕松研獲10+ SCI
-
07.01
“稻”亦有道——盤點(diǎn)近期水稻研究的重大突破
-
06.28
科學(xué)與美學(xué)的結(jié)合體:植物亞細(xì)胞定位技術(shù)詳解
-
06.28
“聚焦新質(zhì)生產(chǎn)力,激發(fā)科研新動(dòng)能”|LCA躋身蛋白互作研究的新銳力量
-
06.05
知無不“研”|一文讀懂免疫共沉淀技術(shù)(Co-IP)
-
05.14
四大研究利器(Co-IP、BIFC、Y2H、GST pull-down)助力速配蛋白互作“最佳拍檔”
-
05.14
高效、精準(zhǔn)、直觀、實(shí)時(shí)——取經(jīng)“蛋白互作研究翹楚”BIFC!
-
05.14
轉(zhuǎn)染效率低、干擾效果差、重復(fù)性欠佳...siRNA研究頻遇“攔路虎”怎么辦?
-
04.22
一文讀懂EMSA技術(shù)核心要點(diǎn),讓“emsa” 秒變“easy”
找回密碼
注冊(cè)帳號(hào)
 X
X -
 在線客服
在線客服 -

 在線訂購
在線訂購 -

 掃碼領(lǐng)資料
掃碼領(lǐng)資料掃碼領(lǐng)資料

電話:19522948401
-

 文獻(xiàn)獎(jiǎng)勵(lì)申請(qǐng)
文獻(xiàn)獎(jiǎng)勵(lì)申請(qǐng)