
IF=15+重慶醫科大學陳教授課題組發現circSEPT9可作為TNBC潛在的生物標志物和治療靶點
信息來源:金開瑞 作者:genecreate 發布時間:2020-07-23 13:02:12
題目:The circRNA circSEPT9 mediated by E2F1 and EIF4A3 facilitates the carcinogenesis and development of triple-negative breast cancer
期刊名稱:Mol. Cancer
影響因子:15.302
主要技術:RNA pull down、雙熒光素酶、RIP、ChIP
研究背景
越來越多的研究表明,circRNA與許多癌癥的發生發展密切相關。然而,環狀RNA在三體陰性乳腺癌(TNBC)中的生物學功能和潛在的分子機制仍不清楚。作者運用高通量RNA測序技術研究了circRNA在四對三陰性乳腺癌及癌旁組織中的表達模式,并通過qPCR和原位雜交技術評估circSEPT9的表達和預后意義,并進行了一系列的體內和體外功能檢測實驗,以研究circSEPT9在三陰性乳腺癌變及發展過程中的調控作用。同時采用Chip、雙熒光素酶報告基因及RIP等檢測方法探討了E2F1和EIF4A3對circSEPT9的潛在調控作用,驗證了circSEPT9和mir-637的互作關系。
技術路線
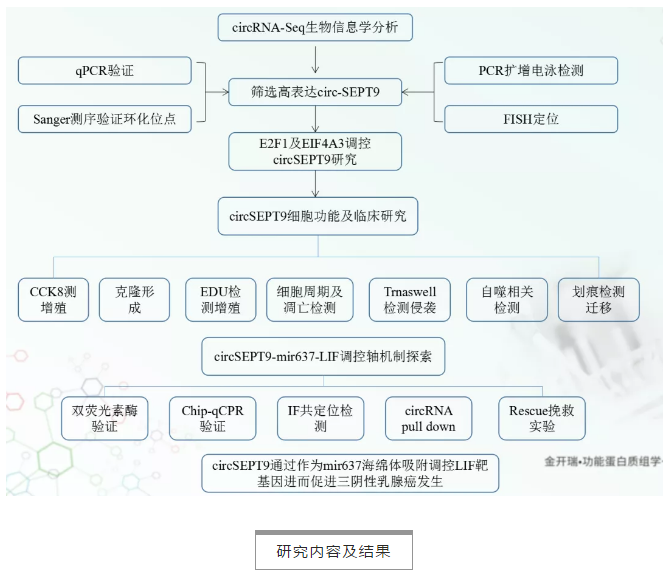
研究內容及結果
1. circRNA測序及生物信息學分析篩選circSEPT9
4對三陰性乳腺癌及癌旁組織,通過circRNA二代測序,根據生物信息學分析發現354個差異表達circRNAs,其中上調circRNA 47個,下調circRNA 354個(篩選條件:fold_change≥2,P-value值<0.05)。作者從TOP30個差異表達circRNA中又進一步篩選到一個novel circRNA,circSEPT9(hsa_circ_0005320,由SEPT9第二號外顯子反向剪切形成)作為項目研究重點(Fig. 1a)。針對circSEPT,作者從以下幾個方面層層進行驗證:
(1). qPCR檢測驗證其在三陰性乳腺癌中表達上調
(2). sanger 測序法驗證circSEPT環化位點(Fig.1b)
(3). 分別設計divergent引物及convergent引物,以cDNA及gDNA為模板分別進行擴增檢測,驗證其確實為環狀RNA(Fig.1c)
(4). 通過擴增及電泳檢測發現circSEPT9相比于線性SRPT9對RNase R消化的抵抗能力更高(Fig. 1d&e)
(5). 使用放線菌素D抑制劑評估circSEPT9的穩定性(Fig. 1f)
(6). FISH實驗對circSEPT9進行定位,主要分布于細胞質中(Fig1g&h)
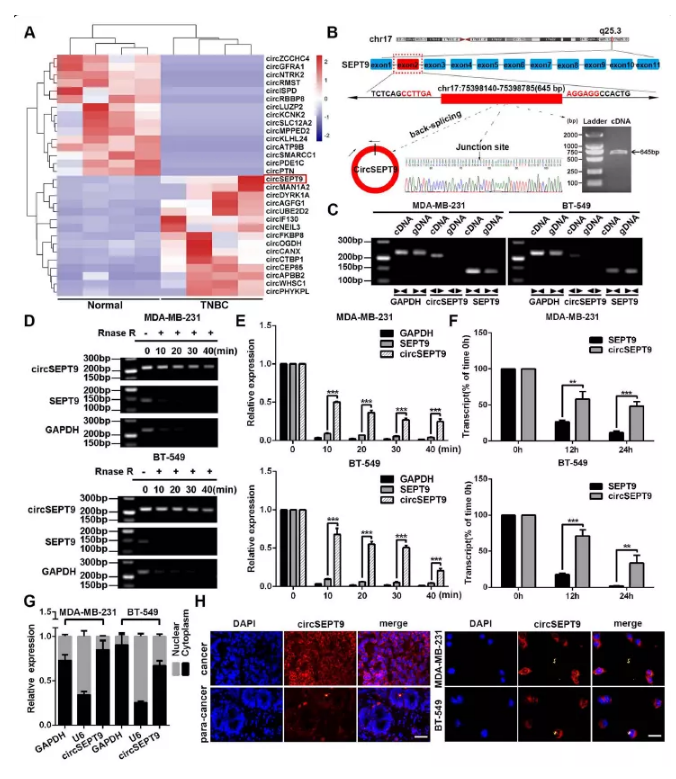
圖1 三陰性乳腺癌細胞中circSEPT9的驗證及檢測
A. 4對三陰性乳腺癌及癌旁樣品top15差異circRNA聚類熱圖,紅色代表上調,藍色代表下調
B. sanger測序驗證環化位點及qPCR檢測表達量
C. 以cDNA及gDNA進行擴增及凝膠電泳檢測
D&E. 以RNase R處理三陰性乳腺癌細胞并通過PCR擴增檢測其表達量情況
F. 放線菌素D處理12及24h后,通過qPCR檢測三陰性乳腺癌細胞中circSEPT9和線性SEPT9的表達
G. 核質分離檢測circSEPT9表達,以GAPDH作為質對照,U6作為核對照
H. FISH檢測實驗觀察circSEPT9定位,分別以組織及細胞進行染色拍照,細胞核采用DAPI染色
2. E2F1和EIF4A3促進circSEPT9的表達
E2F1是E2F轉錄因子家族成員,可在轉錄水平調控靶基因的表達。作者通過生物信息學網站預測(http://jaspar.genereg.net)發現SEPT9啟動子上有13個與E2F1結合的位點,作者從中選擇了3個最接近的結合位點,通過雙熒光素酶素酶及chip-qPCR進行驗證。為探索E2F1和EIF4A3對circSEPT9表達的影響,分別構建過表達質粒,同時合成干擾siRNA,轉染三陰性乳腺癌細胞進行過表達及干擾效果檢測,后進行qPCR檢測circSEPT9表達變化。
實驗結果如下:
(1). 篩選驗證有效的過表達質粒及干擾siRNA分別轉染三陰性乳腺癌細胞MDA-MB-231及BT-549細胞后,qPCR檢測SEPT9表達,結果顯示:E2F1促進SEPT9的表達(Fig. 2a)
(2). 通過構建野生型及突變型載體,通過雙熒光素酶實驗檢測生信預測SEPT9啟動子結合位點,結果顯示:所篩選2號及3號預測結合位置與E2F1結合,1號不結合(Fig. 2b&c)
(3). 利用E2F1對應Chip級別抗體進行染色質免疫共沉淀實驗,實驗獲取互作DNA產物進行Chip-qPCR實驗分析結果表明:E2F1可以結合SEPT9基因啟動子區并加速其轉錄活性(Fig2d)
小結:以上部分數據證明SEPT9是轉錄因子E2F1的靶基因。
EIF4A3是外顯子連接復合體的核心組成部分,在mRNA前期剪接中起著至關重要的作用。作者接下來又從circSEPT9的上游和下游區域找到5個與EIF4A3結合位點,并通過RIP實驗驗證其互作位點信息。同時在過表達及干擾EIF4A3后qPCR檢測circSEPT9表達水平變化。作者進一步通過敲低circSEPT9后檢測三陰性乳腺癌細胞中細胞周期相關蛋白的表達變化,并通過挽救實驗共轉過表達質粒后再次檢測circSEPT9表達變化,實驗結果如下:
(1). E2F1過表達及干擾siRNA分別轉染MDA-MB-231及BT-549細胞后,qPCR檢測circSEPT9表達量變化,結果顯示:E2F1上調顯著促進了circSEPT9的表達,干擾E2F1表達后抑制circSEPT9的表達(Fig.2e)
(2). RIP分析結果顯示:EIF4A3可以通過推定的結合位點與SEPT9 pre-mRNA結合(Fig. 2g)
(3). qPCR結果顯示:過表達EIF4A3促進circSEPT9的表達,干擾EIF4A3后抑制circSEPT9表達(Fig. 2h)
(4). 干擾circSEPT9后,細胞周期相關蛋白表達下降(Fig.2i)
小結:以上數據顯示EIF4A3通過促進circSEPT9的表達進而調節細胞周期相關蛋白的表達。
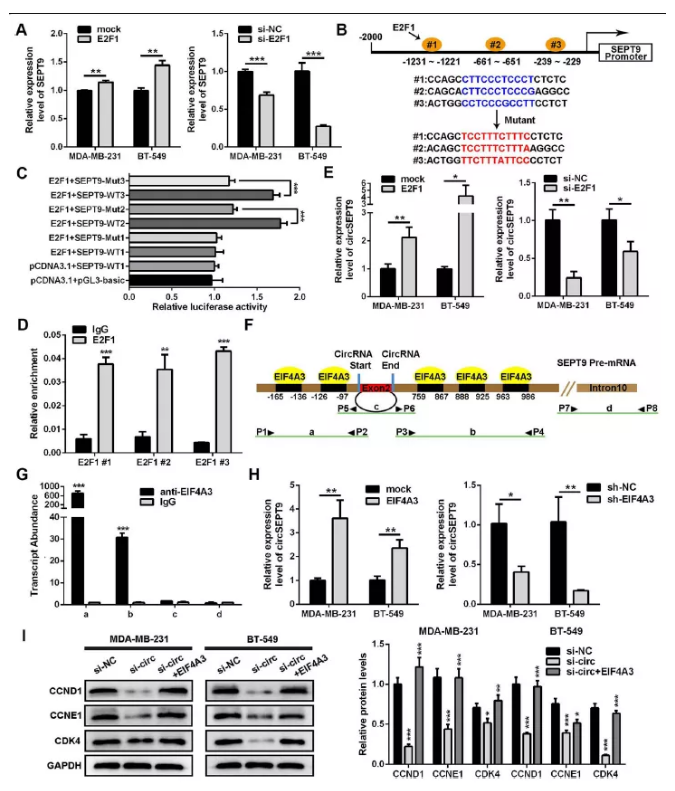
圖2 E2F1和EIF4A3分別與SEPT9啟動子和pre-mRNA結合增強circSEPT9的表達
(A) qPCR檢測過表達及干擾E2F1后三陰性乳腺癌細胞中SEPT9表達變化。
(B) 生物信息學預測并選定的SEPT9啟動子上E2F1 3個結合位點,其中包括野生型(WT)及突變型(Mut)序列。
(C) 雙熒光素酶檢測SEPT9與E2F1對應3個結合位點。
(D) 在MDA-MB-231細胞中進行Chip-qPCR檢測驗證結合位點
(E) E2F1過表達及干擾siRNA轉染三陰性乳腺癌細胞后qPCR檢測circSEPT9表達變化
(F&G)RIP實驗證實EIF4A3可以與SEPT9 pre-mRNA結合
(H)過表達及干擾EIF4A3后,qPCR檢測circSEPT9表達
(I)WB檢測細胞周期相關蛋白表達及灰度值統計(*P<0.05,**P<0.01,***P<0.001)
3. 細胞及臨床研究證實circSEPT9高表達
臨床收集60對三陰性乳腺癌及癌旁組織樣品,同時在三陰性乳腺癌細胞MDA-MB-231、BT-549、MDA-MB-453、MDA-MB-468及正常乳腺上皮細胞MCF-10A中進行qPCR檢測circSEPT9表達,結果顯示circSEPT9在三陰性乳腺癌細胞及組織中的表達均高于正常乳腺上皮細胞及癌旁組織,與RNA-Seq結果保持一致(Fig. 3a&b)。作者進一步繪制ROS曲線用于評估circSEPT9對三陰性乳腺癌篩查的診斷價值,結果顯示:circSEPT9的曲線下面積(AUC)為0.711,在截距為1.971時,特異性和敏感性分別為75%和63.3%(Fig. 3c)。Kaplan-Meier生存分析結果顯示:乳腺癌患者中circSEPT9高表達(P=0.001)。N2-3階段(P=0.001)和TNM II/III期(P=0.031)總體生存率低于N0-1期和TNM I期(Fig. 3g),預示著circSEPT9可以作為腫瘤標記物,高表達circSEPT9造成患者的不良預后。
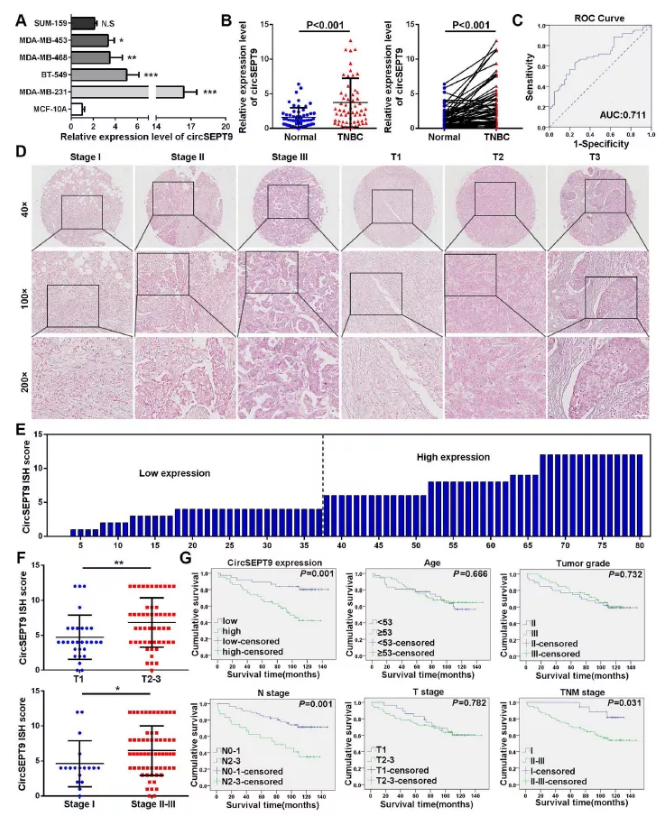
圖3 circSEPT9在三陰性乳腺癌患者中高表達并與患者不良預后有關
(A和B) qPCR檢測60對臨床三陰性乳腺癌及癌旁樣品及三陰性乳腺癌細胞及正常乳腺上皮細胞中circSEPT9表達
(C)ROC曲線評估
(D)原位雜交檢測circSEPT9在80個乳腺癌組織中的表達(比例尺,50um)
(E)在80個TNBC組織中計算ISH染色分數,ISH染色得分<6被定義為低表達,≥6視為高表達
(F)在80例不同的三陰性乳腺癌患者中顯示circSEPT9 IHC評分的點分布圖
(G) Kaplan-Meier生存曲線分析圖
4. circSEPT9促進三陰性乳腺癌細胞的增殖、遷移和侵襲
為探索circSEPT9潛在生物學功能,作者構建了circSEPT9過表達載體及干擾siRNA(Fig. 4a),分別轉染細胞后qPCR檢測確定其過表達及干擾效果(Fig. 4b&c),同時用線性SEPT9引物進行檢測發現過表達及敲低circSEPT9后對線性SEPT9基因的表達水平沒有顯著變化(Fig. 4d)。經克隆形成,CCK8及EDU分析其細胞生物表現變化,結果顯示:
(1). 干擾circSEPT9顯著抑制了三陰性乳腺癌細胞的增殖能力(Fig. 4e-g)
(2). 過表達circSEPT9能有效增強細胞活力
(3). 干擾circSEPT9抑制了細胞的遷移和侵襲能力。過表達后相反(Fig. 4h-j)
小結:circSEPT9在三陰性乳腺癌細胞中發揮致癌作用
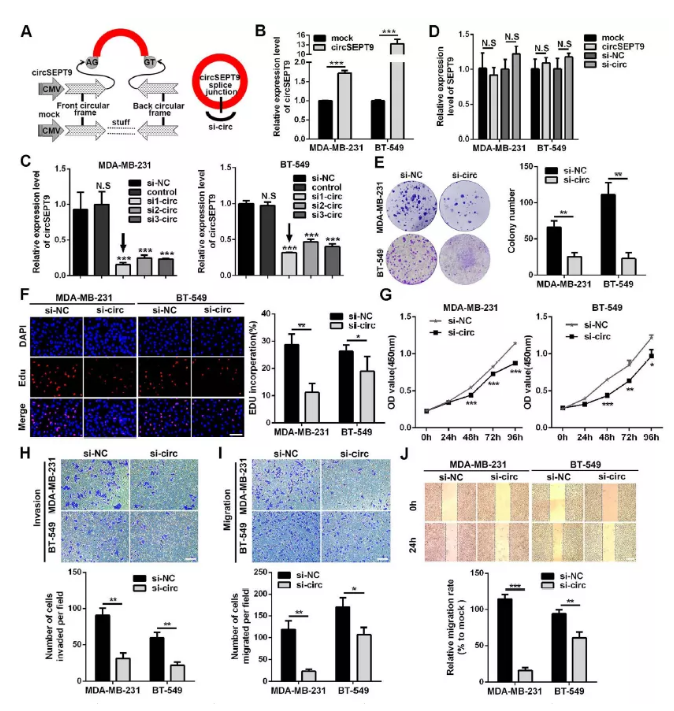
圖4 干擾cricSEPT9降低三陰性乳腺癌細胞的生長、遷移和侵襲能力
(A)circSEPT9過表達載體及si-circSEPT9示意圖
(B-D)qPCR檢測三陰性乳腺癌細胞中circSEPT9過表達及干擾效率
(E) 克隆形成檢測細胞增殖能力
(F) EDU檢測細胞增殖能力
(G) CCK8檢測細胞生長曲線
(H) Transwell檢測細胞侵襲能力(×100,100um)
(I&J) 劃痕實驗檢測細胞遷移能力 (*P<0.05,**P<0.01,***P<0.001)
5. circSEPT9下調誘導細胞周期停滯、引發細胞的凋亡和自噬
作者在干擾circSEPT9的基礎上,進一步進行了三陰性乳腺癌細胞細胞周期,凋亡和自噬等方面的檢測。結果如下:
(1). 細胞周期檢測結果顯示:干擾circSEPT9后,細胞多分布于G1期,少數分布于S期(Fig. 5a)
(2). Western-Blot實驗結果顯示干擾circSEPT9后,細胞周期相關蛋白表達下降(Fig. 5b)
(3). 流式細胞術檢測凋亡實驗結果顯示:下調circSEPT9后,細胞凋亡率顯著升高(Fig. 5c)
(4). TUNEL分析結果進一步證實了流式的結果(Fig. 5d)
(5). Hoechst33342染色實驗結果發現干擾circSEPT9后乳腺癌細胞出現典型核收縮,核破裂等凋亡等特征(Fig. 5e)
(6). WB檢測凋亡相關蛋白結果顯示:下調circSEPT9后,凋亡相關蛋白caspase-3和凋亡蛋白Bax的表達量增加,Bcl-2的表達水平降低
(7). 通過IF免疫熒光檢測顯示自噬相關指標LC3-II在干擾circSEPT9組中增加(Fig. 5g),自噬相關蛋白ATG5和ATG7顯著增加(Fig. 5h)
小結:circSEPT9參與調節細胞周期,凋亡及自噬等相關過程
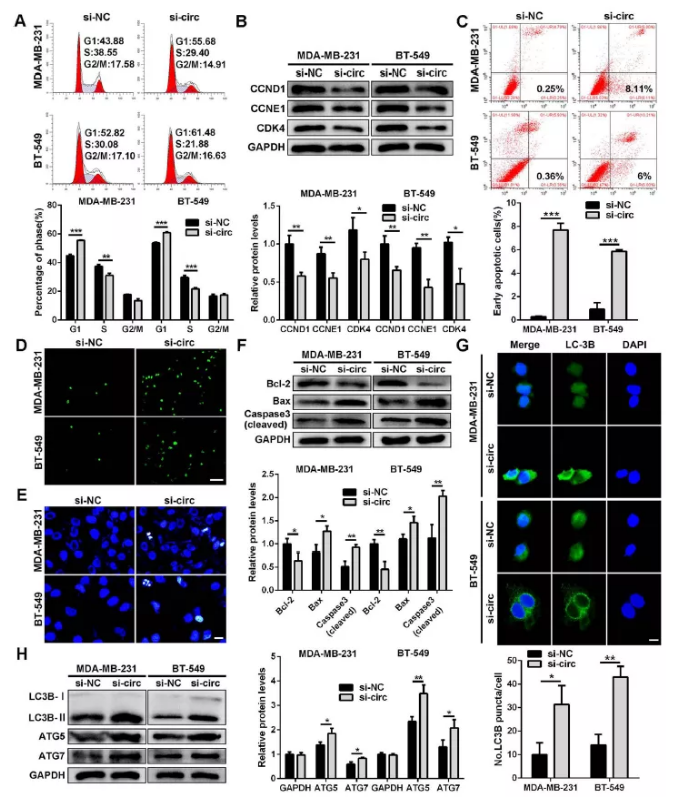
圖5 干擾circSEPT9誘導三陰性乳腺癌細胞周期停滯,細胞凋亡和自噬
(A)干擾circSEPT9后流式檢測細胞周期
(B)干擾circSEPT9后,WB檢測三陰性乳腺癌細胞中細胞周期相關蛋白表達
(C)干擾circSEPT9后,流式細胞細胞術檢測細胞凋亡
(D)干擾circSEPT9后,TUNEL染色觀察細胞凋亡
(E)干擾circSEPT9后,Hoechst33342染色觀察細胞凋亡形態學特征
(F)干擾circSEPT9后,WB檢測凋亡相關蛋白表達水平
(G) IF免疫熒光檢測LC3-3B熒光強度
(H) WB檢測自噬相關蛋白表達水平(*P<0.05,**P<0.01,***P<0.001)
6. circSEPT9與mir-637直接互作并抑制其活性
為探究circSEPT9潛在分子機制,作者通過生物信息學分析預測其可能結合miRNA數據信息,最終確認mir-637作為其互作靶位點。通過qPCR檢測過表達及干擾circSEPT9后對應mir-637的表達,并采用雙熒光素酶進行互作驗證,FISH檢測其定位情況,并經circRNA pull dwon進一步驗證,結果匯總如下:
(1). 過表達circSEPT9后,mir-637表達量下降,干擾組則相反(Fig. 6a)
(2). 60例臨床樣品檢測mir-637表達,結果顯示其在正常組織中高表達(Fig. 6b)
(3). 雙熒光素酶實驗結果顯示,添加mir-637 mimics后circSEPT9野生型組中熒光素酶報告基因的活性降低,突變組無變化,說明circSEPT9與mir-637直接結合(Fig.6e)
(4). 通過FISH共定位檢測發現二者均定位在細胞質中
(5). 進一步通過構建生物素標記的circSEPT9探針進行circRNA Pull down實驗拉取互作RNA,并通過qPCR檢測miR-637表達變化,進一步證實二者互作結合(Fig. 6h)
小結:circSEPT9與mir-637互作
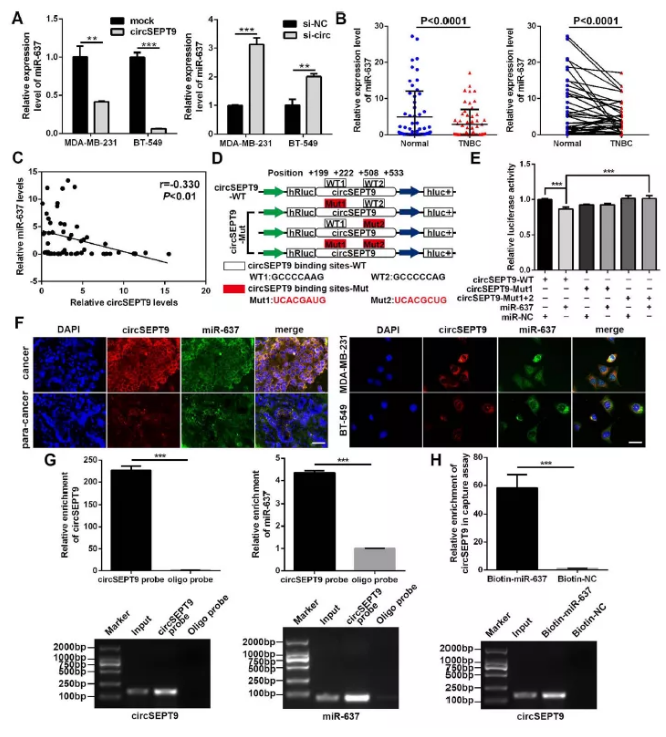
圖6 circSEPT9充當mir-637海綿
(A)過表達及干擾circSEPT9后,qPCR檢測mir-637
(B) 60例臨床三陰性乳腺癌及對照樣品中檢測mir-637
(C) 皮爾遜相關分析
(D) 雙熒光素酶預測結合位點示意圖
(E) 雙熒光素酶檢測實驗
(F) circSEPT9及mir-637共定位檢測
(G) circRNA pull down檢測實驗
7. mir-637與circSEPT9共轉染功能研究
為進一步研究circSEPT9與mir-637互作功能的相關作用,作者在三陰性乳腺癌細胞中進行共轉染實驗,結果顯示:
(1). mir-637過表達基礎上進行circSEPT9過表達后顯著減弱了細胞增殖、遷移及細胞侵襲能力
(2). 在干擾circSEPT9的基礎上進一步干擾mir-637后能逆轉circSEPT9造成的表型變化
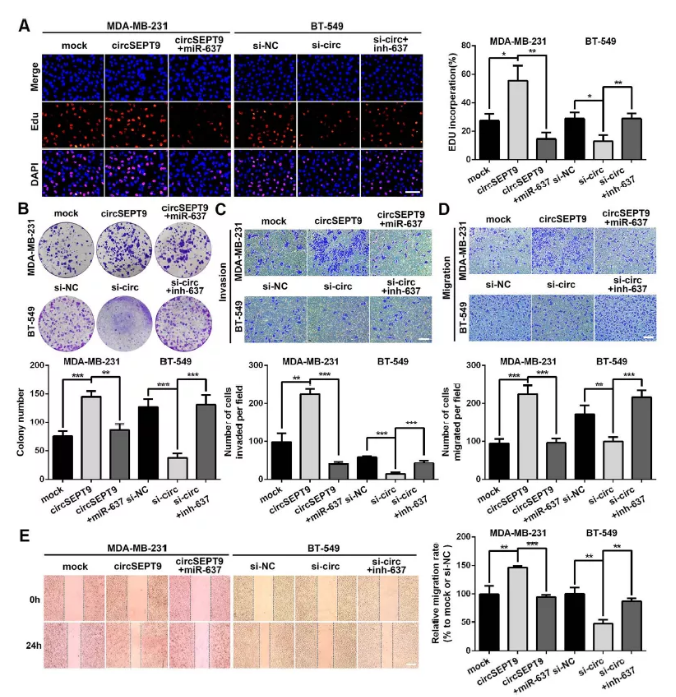
Fig. 7 mir-637可以部分逆轉circSEPT9對三陰性乳腺癌細胞的增殖及遷移作用
(A&B) EDU檢測細胞增殖
(C-E)三陰性乳腺癌細胞中的侵襲和遷移檢測
8. LIF確認為mir-637靶基因激活LIF-STAT3通路發揮作用
為進一步確認確認調控路徑,作者通過Targetscan(http://www.targetscan.org),miRanda(http:// www.microrna.org)以及Findtar在線程序(http://bio.sz.tsinghua.edu.cn),最終確認LIF作為mir-637下游保守靶位點。作者通過一系列實驗對篩選下游靶基因LIF進行檢測,結果如下:
(1). qPCR及wb檢測結果顯示:在過表達mir-637后,LIF表達量下降,干擾后則相反(Fig. 8a&b)
(2). 雙熒光素酶檢測實驗對mir-637及LIF結合情況進行驗證,結果顯示:mir-637 mimics轉染組可以顯著降低LIF野生型組中的熒光值,突變組中無明顯變化(Fig. 8c&d)
(3). 免疫組化IHC及IF免疫熒光檢測顯示:LIF在三陰性乳腺癌組織上調(Fig. 8e&f)
(4). 皮爾遜相關分析表達LIF的表達與mir-637負相關(Fig.8h)
(5). 相關分析表明LIF的表達與circSEPT9呈正相關(Fig. 8i)
(6). qPCR及IF分析表明過表達circSEPT9后,LIF表達量顯著增加,干擾后則下降(Fig. 8k&l)
(7). 進一步結果顯示:過表達circSEPT9上調LIF,p-STAT3、ID1及MDM2,使P53及P21表達下降,干擾后發揮相反的作用。
小結:circSEPT9可以作為mir-637的海綿激活LIF-STAT3途徑促進三陰性乳腺癌變過程
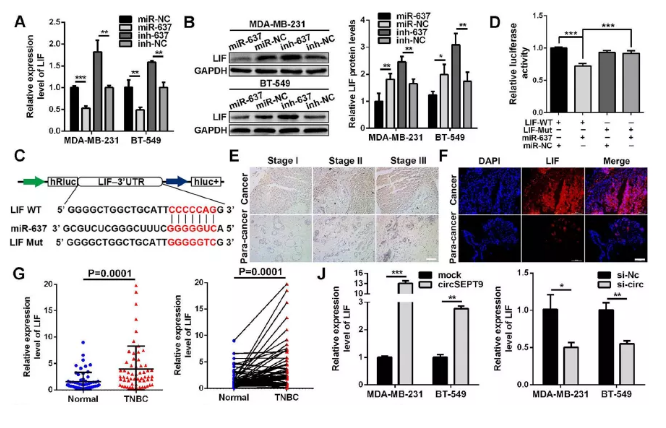
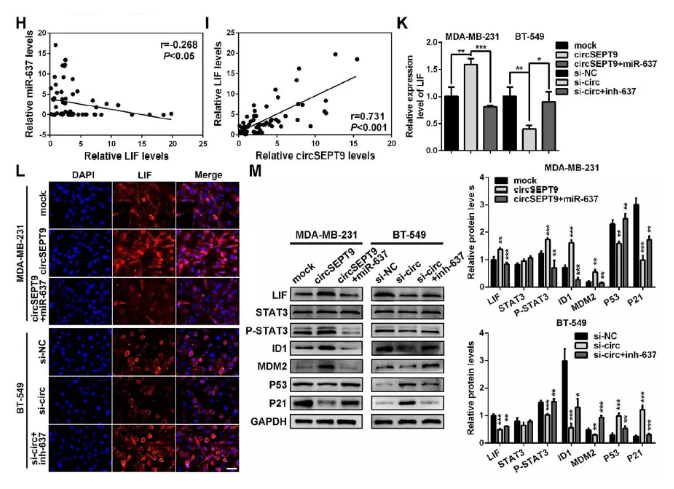
Fig. 8 circSEPT9可以作為mir-637的海綿激活LIF-STAT3途徑促進三陰性乳腺癌變過程
(A&B)qPCR及WB檢測mir-637過表達及干擾后LIF變化
(C&D)雙熒光素酶檢測驗證mir-637與LIF互作結合
(E&F) IHC及IF評估LIF的相對表達
(G)臨床60例三陰性乳腺癌及癌旁樣本中qPCR檢測LIF
(H&I) 臨床樣品中mir-637,circSEPT9與LIF相關分析
(J)qPCR檢測過表達及干擾circSEPT9后LIF表達
(K)共轉染后LIF表達檢測
(L) IF檢測共轉染后細胞中LIF表達
(M)LIF及下游LIF-STAT3途徑相關蛋白表達檢測
小結
作者從circRNA測序表達譜中專注于circSEPT9的機制研究,分別從生物學功能及機制方面對其在三陰性乳腺癌發生發展過程中的調控機制進行研究,最終確認circSEPT9可以作為新的診斷和預后或治療標記物。
最新動態
-
09.23
中藥的現代詮釋:外泌體如何革新傳統醫學?
-
07.02
1+1>2!深度解析RNA測序數據挖掘邏輯和后期實驗設計思路,輕松研獲10+ SCI
-
07.01
“稻”亦有道——盤點近期水稻研究的重大突破
-
06.28
科學與美學的結合體:植物亞細胞定位技術詳解
-
06.28
“聚焦新質生產力,激發科研新動能”|LCA躋身蛋白互作研究的新銳力量
-
06.05
知無不“研”|一文讀懂免疫共沉淀技術(Co-IP)
-
05.14
四大研究利器(Co-IP、BIFC、Y2H、GST pull-down)助力速配蛋白互作“最佳拍檔”
-
05.14
高效、精準、直觀、實時——取經“蛋白互作研究翹楚”BIFC!
-
05.14
轉染效率低、干擾效果差、重復性欠佳...siRNA研究頻遇“攔路虎”怎么辦?
-
04.22
一文讀懂EMSA技術核心要點,讓“emsa” 秒變“easy”
 X
X 






