
iTRAQ蛋白質(zhì)組學(xué)揭示油菜卷葉機(jī)理
信息來源:金開瑞 作者:genecreate 發(fā)布時(shí)間:2018-06-21 11:03:06
題目:Histological, Physiological, and Comparative Proteomic Analyses Provide Insights into Leaf Rolling in Brassica napus
期刊:Journal of Proteome Research
影響因子:4.268
合作技術(shù):iTRAQ
研究背景
葉片是大多數(shù)植物光合作用的主要器官,葉片發(fā)育影響作物產(chǎn)量和植物結(jié)構(gòu)。適度的卷葉被認(rèn)為是作物理想型育種的一個(gè)重要組成部分,它能夠改變植物的結(jié)構(gòu),提高光合效率,延緩葉片衰老,減輕干旱、高溫和高光等脅迫帶來的損害。近年來,在擬南芥和水稻等物種中,已經(jīng)分離出了卷曲葉片突變體,并對(duì)它們的分子機(jī)制進(jìn)行了深入的研究。油菜作為世界上最重要的油料作物之一,雖然也發(fā)現(xiàn)了一些卷曲葉片的突變體,但是對(duì)于其作用機(jī)制仍有待進(jìn)一步的研究。
本文作者主要利用iTRAQ定量蛋白質(zhì)組學(xué)技術(shù)分析油菜卷葉突變株Bndcl1與野生株(WT)的差異蛋白,闡明了卷葉機(jī)理及其對(duì)植物的生理影響,有望促進(jìn)油菜的理想株型育種。
研究?jī)?nèi)容及結(jié)果
1. 本研究中,作者選取了油菜卷葉突變株Bndcl1,首先經(jīng)組織學(xué)對(duì)比發(fā)現(xiàn),相比于野生株(WT),突變株(Bndcl1)幼苗期的葉片表型上比較緊湊(圖 1A和B)。組織切片結(jié)果中發(fā)現(xiàn)Bndcl1葉綠體數(shù)目明顯增多,特別是在海綿狀的葉肉細(xì)胞中(圖1C和D)。此外,作者發(fā)現(xiàn),在Bndcl1突變體中,在遠(yuǎn)軸端韌皮部組織細(xì)胞較少,而在其葉脈以下的遠(yuǎn)軸端表皮細(xì)胞明顯增大(圖1E和F)。
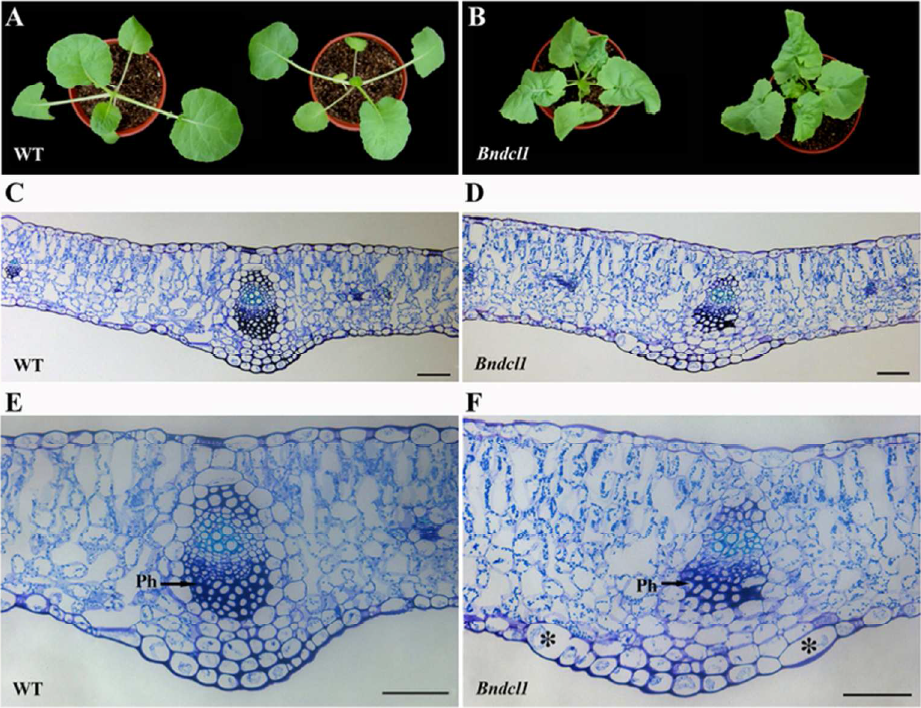
圖1.葉片的組織學(xué)分析
2. 為了評(píng)估卷曲葉片對(duì)Bndcl1突變株光合性能的影響,作者測(cè)定了Chl的含量、Chl熒光和氣體交換參數(shù)。與野生株相比,Bndcl1株中的葉綠體數(shù)量、PSII和凈光合速率的有效量子產(chǎn)量顯著增加。
3. 活性氧(ROS)的生成在光合作用的光反應(yīng)過程中是不可避免的,PSII極易受到光破壞的影響。光抑制的光合作用不可避免的損失會(huì)因ROS的產(chǎn)生而加劇,而ROS的產(chǎn)生會(huì)減慢PSII的修復(fù)。為了減少光氧化傷害,光合生物體已經(jīng)進(jìn)化出多種機(jī)制,而對(duì)抗氧化劑的上調(diào)是調(diào)節(jié)機(jī)制的一部分。因此,作者檢測(cè)了Bndcl1和WT葉子的酶活性、脂質(zhì)過氧化和ROS水平,發(fā)現(xiàn)在Bndcl1葉片中O2• − 和H2O2較低,由此說明其ROS水平也將明顯降低(圖2A和B)。此外,TBARS在突變株的低水平同樣說明其膜脂過氧化的水平降低(圖2C)。作者還檢測(cè)比較了兩者的SOD、POD和CAT酶活性,發(fā)現(xiàn)相比于WT株,在Bndcl1葉片中此酶活性顯著增加。由此說明Bndcl1的光系統(tǒng)能更好的避免受到光氧化傷害。
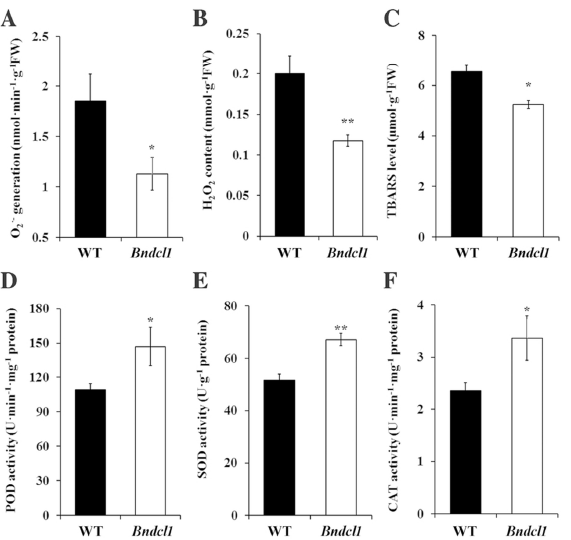
圖2. 野生株(WT)和突變株(Bndcl1)的葉子酶活性、脂質(zhì)過氧化和ROS水平
4. 作者應(yīng)用iTRAQ定量蛋白質(zhì)組學(xué)技術(shù)研究Bndcl1與WT葉片蛋白質(zhì)表達(dá)水平的差異,總共鑒定到5019個(gè)蛋白,差異蛋白有943個(gè),其中發(fā)生上調(diào)的蛋白有451個(gè),發(fā)生下調(diào)的蛋白有492個(gè)。作者對(duì)這些蛋白進(jìn)行了GO、COG、KEGG等注釋分析。通過對(duì)差異蛋白中發(fā)生上調(diào)和下調(diào)的蛋白分別進(jìn)行GO分析并對(duì)比發(fā)現(xiàn),在“細(xì)胞增殖”、“生物過程的調(diào)節(jié)”、“色素沉著”等功能方面,下調(diào)的蛋白數(shù)目要比上調(diào)的蛋白數(shù)多,具有抗氧化活性的功能的上調(diào)蛋白數(shù)目是下調(diào)蛋白數(shù)目的3倍多(圖3)。COG分析表明,大多數(shù)下調(diào)蛋白參與翻譯、核糖體結(jié)構(gòu)和生物發(fā)生,而上調(diào)蛋白主要參與碳水化合物的運(yùn)輸和代謝、轉(zhuǎn)錄翻譯后修飾、蛋白質(zhì)的周轉(zhuǎn)、分子伴侶和能量的產(chǎn)生和轉(zhuǎn)化(圖4)。KEGG 注釋結(jié)果顯示,盡管在“代謝途徑”、“次生代謝產(chǎn)物的生物合成”和“淀粉和蔗糖代謝”中均有上調(diào)和下調(diào)的蛋白質(zhì)參與,但在不同的途徑中也涉及到不同差異蛋白質(zhì)(圖5)。
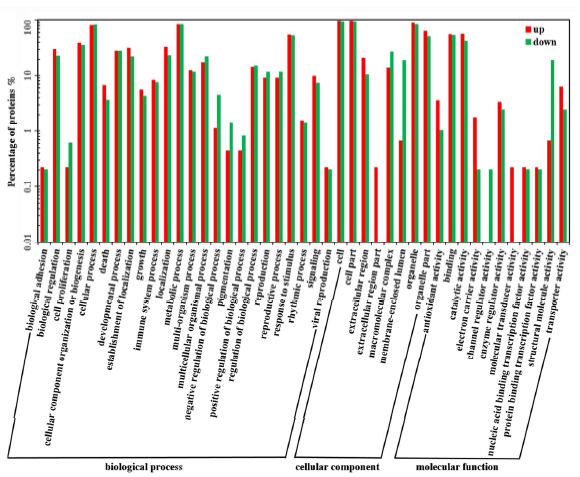
圖3.差異表達(dá)蛋白的GO功能注釋分析
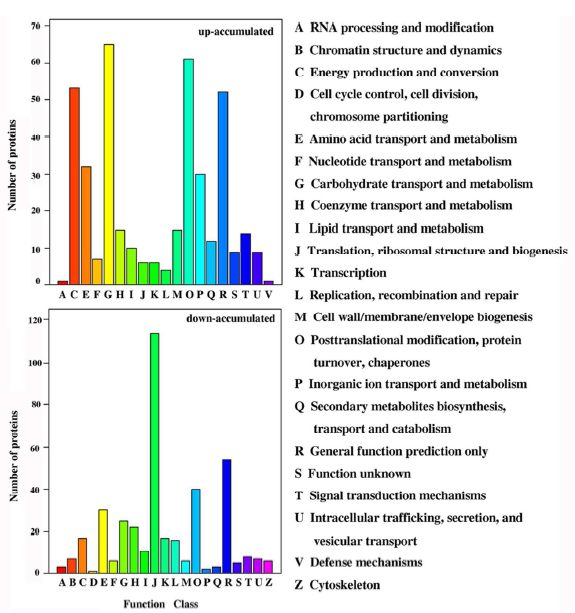
圖4. 差異表達(dá)蛋白的COG注釋分析
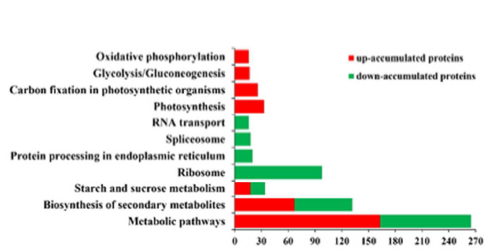
圖5. 差異表達(dá)蛋白的KEGG富集分析
5. 為了進(jìn)一步對(duì)這些差異蛋白的功能進(jìn)行分類分析,作者進(jìn)行了功能富集分析。通過對(duì)差異蛋白的富集分析結(jié)果發(fā)現(xiàn),這些發(fā)生上調(diào)的蛋白能夠改善Bndcl1株的光合性能和減輕氧化傷害,并且發(fā)現(xiàn)下調(diào)的蛋白參與了葉片近-遠(yuǎn)軸極性建成。
6. 作者最后從差異表達(dá)蛋白中挑選了多個(gè)蛋白進(jìn)行了qRT-PCR表達(dá)驗(yàn)證。例如:與卷葉表型相關(guān)的基因PGY1, PGY2, PGY3, AE5, STV1 and SGS3,這幾個(gè)基因的蛋白表達(dá)水平在蛋白質(zhì)組學(xué)結(jié)果是顯著下調(diào)。qRT-PCR驗(yàn)證結(jié)果顯示,在Bndcl1株,這些基因mRNA表達(dá)水平顯著降低,表明這些基因在蛋白質(zhì)和mRNA水平上發(fā)生了一致的變化(圖6A)。另外作者從轉(zhuǎn)錄水平上檢測(cè)了葉極性的關(guān)鍵調(diào)節(jié)因子:AS1, AS2, KAN1, KAN2, REV, RDR6, AGO7, AGO1(圖6B)。結(jié)果發(fā)現(xiàn)在Bndcl1株,AS1, KAN2發(fā)生了顯著降低,AS2, KAN1沒有顯著變化,REV發(fā)生了顯著下調(diào)。此外,RDR6, AGO7, AGO1也同樣發(fā)生了顯著下調(diào)。最后作者還對(duì)油菜甾醇相關(guān)的6個(gè)差異蛋白進(jìn)行了轉(zhuǎn)錄水平的驗(yàn)證,結(jié)果與iTRAQ數(shù)據(jù)(圖6C)一致,說明了定量蛋白質(zhì)組學(xué)分析所獲得的結(jié)果的可靠性。
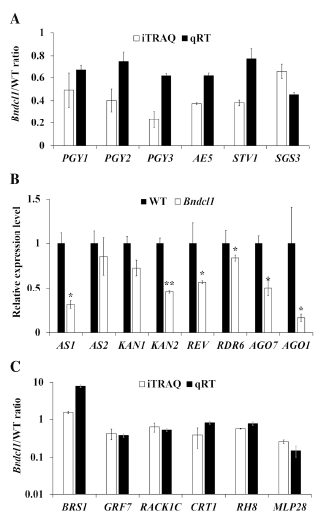
圖6. qRT-PCR驗(yàn)證分析
文章小結(jié)
這項(xiàng)結(jié)果研究表明,葉片近-遠(yuǎn)軸極性和油菜甾醇代謝及信號(hào)通路的缺陷是導(dǎo)致Bndcl1突變株葉子向下卷曲表型的原因,葉片的適度卷曲能夠提高光吸收、能量轉(zhuǎn)移和CO2固定效率,進(jìn)而提高了突變株的光合性能。此外,在突變株中,參與PSII修復(fù)周期的抗氧化劑和蛋白質(zhì)水平的升高可能會(huì)減少非生物脅迫的光合損失。這項(xiàng)研究為油菜卷葉機(jī)制及卷葉對(duì)植物生理的影響提供了新的見解。因此,卷葉突變株因具有緊湊的結(jié)構(gòu),能提高光合效率和增強(qiáng)抗逆性,這也是作物育種理想株型。
解析文獻(xiàn)
Wenjing Chen, Shubei Wan, et al. Histological, Physiological, and Comparative Proteomic Analyses Provide Insights into Leaf Rolling in Brassica napus. 2018, DOI: 10.1021/acs.jproteome.7b00744
參考文獻(xiàn)
1. Yang, C.; Xu, L.; et al. iTRAQ-based proteomics of sunflower cultivars differing in resistance to parasitic weed Orobanche cumana. Proteomics 2017, 17, 1700009.
2. Liang, J.; Liu, B.; et al. Genetic variation and divergence of genes involved in leaf adaxial-abaxial polarity establishment in Brassica rapa. Front. Plant Sci. 2016, 7, 94.
3. Husbands, A. Y.; Benkovics, A. H.; et al. The ASYMMETRIC LEAVES complex employs multiple modes of regulation to affect adaxial-abaxial patterning and leaf complexity. Plant Cell 2015, 27, 3321-3335.
4. Jia, H.; Shao, M.; et al. Proteome dynamics and physiological responses to short-term salt stress in brassica napus leaves. Plos One 2015, 10, e0144808.
5. Yamaguchi, T.; Nukazuka, A.; Tsukaya, H. Leaf adaxial-abaxial polarity specification and lamina
outgrowth: Evolution and development. Plant Cell Physiol. 2012, 53, 1180-1194.
最新動(dòng)態(tài)
-
09.23
中藥的現(xiàn)代詮釋:外泌體如何革新傳統(tǒng)醫(yī)學(xué)?
-
07.02
1+1>2!深度解析RNA測(cè)序數(shù)據(jù)挖掘邏輯和后期實(shí)驗(yàn)設(shè)計(jì)思路,輕松研獲10+ SCI
-
07.01
“稻”亦有道——盤點(diǎn)近期水稻研究的重大突破
-
06.28
科學(xué)與美學(xué)的結(jié)合體:植物亞細(xì)胞定位技術(shù)詳解
-
06.28
“聚焦新質(zhì)生產(chǎn)力,激發(fā)科研新動(dòng)能”|LCA躋身蛋白互作研究的新銳力量
-
06.05
知無不“研”|一文讀懂免疫共沉淀技術(shù)(Co-IP)
-
05.14
四大研究利器(Co-IP、BIFC、Y2H、GST pull-down)助力速配蛋白互作“最佳拍檔”
-
05.14
高效、精準(zhǔn)、直觀、實(shí)時(shí)——取經(jīng)“蛋白互作研究翹楚”BIFC!
-
05.14
轉(zhuǎn)染效率低、干擾效果差、重復(fù)性欠佳...siRNA研究頻遇“攔路虎”怎么辦?
-
04.22
一文讀懂EMSA技術(shù)核心要點(diǎn),讓“emsa” 秒變“easy”
找回密碼
注冊(cè)帳號(hào)
 X
X -
 在線客服
在線客服 -

 在線訂購(gòu)
在線訂購(gòu) -

 掃碼領(lǐng)資料
掃碼領(lǐng)資料掃碼領(lǐng)資料

電話:19522948401
-

 文獻(xiàn)獎(jiǎng)勵(lì)申請(qǐng)
文獻(xiàn)獎(jiǎng)勵(lì)申請(qǐng)