
三維基因組學經典好文:LHX2-和LDB1的反式相互作用調節嗅覺受體的選擇
信息來源:金開瑞 作者:genecreate 發布時間:2019-02-28 08:59:51
題目:LHX2- and LDB1-mediated trans interactions regulate olfactory receptor choice
LHX2-和LDB1的反式相互作用調節嗅覺受體的選擇
期刊:Nature
影響因子:41.577
研究背景
我們的鼻子可以幫助我們感知到玫瑰花香、茉莉花香等各種不同的味道,是什么保證了我們能擁有這嗅覺了?哥倫比亞大學的研究人員借助Hi-C基因組測序技術,首次在哺乳動物的細胞核內發現一種巧妙的三維重排機制。
文章思路
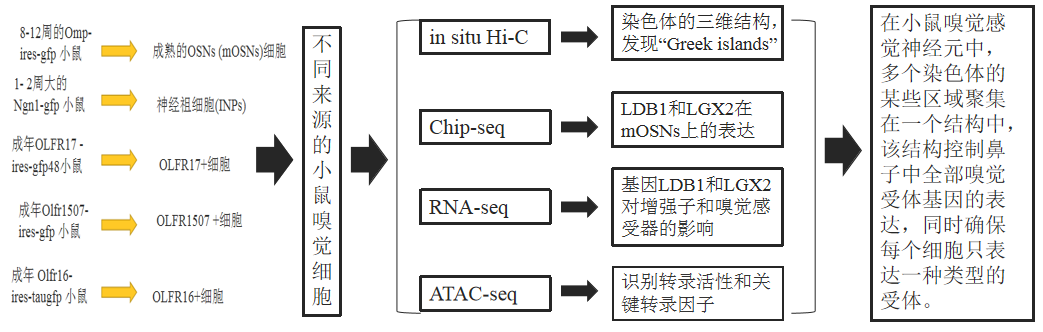
名詞解釋:
小鼠嗅覺感受器(ORs)(olfactory receptors)
成熟嗅覺神經元(mOSN)(mature olfactory sensory neuron)
嗅覺神經元(OSN)(olfactory sensory neuron)
主嗅上皮(MOE)(main olfactory epithelium)
熒光活化細胞(FAC)(fluorescence-activated cell)
總峰分析(APA)(Aggregate peak analysis)
水平基底細胞(HBCs)(horizontal basal cells)
直接神經元前體(INPs)(immediate neuronal precursors)
細菌假染色體(BAC)bacterial artificial chromosome
研究內容及結果
1. 反式分隔來源于OSN分化
首先,作者分析了熒光活化細胞(FAC)分化的成熟嗅覺神經元(mOSNs),它代表了有絲分裂后的神經元的最終分化,其被視為小鼠嗅覺感受器(OR)的異質性。在成熟嗅覺神經元的原位Hi-C結果中顯示了廣泛的染色體間相互作用,相當于Hi-C總接觸的35.6%,其與之前的成像結果一致。放大的基因組視圖顯示,在OR基因簇與中間的OR簇之間存在很強的OSN-specific反式接觸,有大約7.5%的Hi-C接觸點從不同的染色體映射到OR簇(圖1a,b)。聚合峰分析(APA)和無偏室預測證實大多數OR簇參與OR選擇性多染色體的組裝(圖1b),值得注意的是,在mOSNs中,OR簇間的反式接觸僅占所有染色體間接觸的0.25%,但在1000個最強的反式Hi-C接觸中占50%。在分化程度更高的直接神經元前體(INPs)中,反式OR接觸豐富,但比在mOSNs少;順式OR簇間的相互作用程度與mOSNs一致。因此,OR基因間隔以分層的方式形成,順式接觸首先出現,反式作用隨著分化而加強。
OR隔間內,63常染色質的Greek islands代表Hi-C“hotspots”中特殊和頻繁的順式和反式接觸(圖1d,e)。在INPs中,每個細胞中有多個OR基因被弱轉錄。雖Greek islands之間相互作用,但在mOSNs中缺乏焦點接觸分布。分化增強和反式交互規范化是大多數Greek islands的特性(圖1 f, g)。總而言之,在mOSNs中,Greek island Hi-C接觸點中4.5%與其他Greek island有接觸,其中一半為反式(圖1f)。值得注意的是,這超過了Greek islands順式長交互中LHX2和EBF共結合基因間序列的平均和累積頻率(圖1g)。
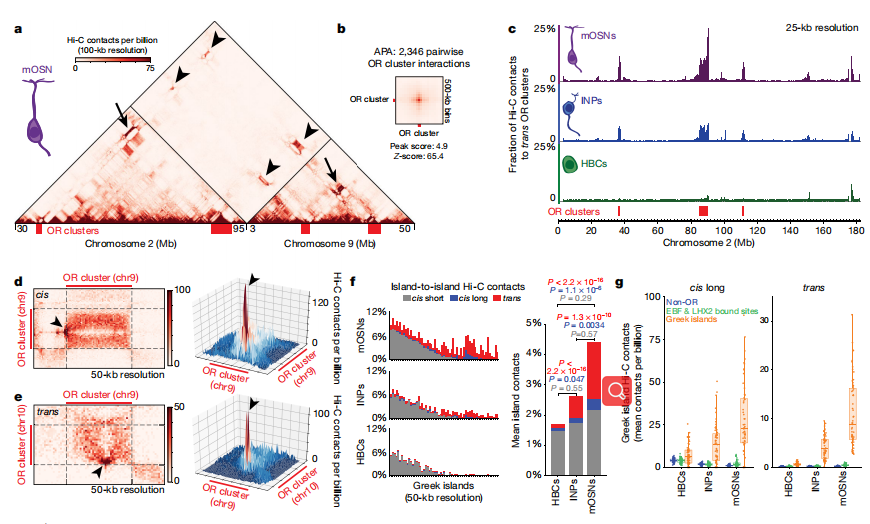
圖1 OSN分化過程中,Greek islands之間形成的染色體間接觸與OR基因簇間接觸以及局部染色體間接觸
2. Greek islands促進區域劃分
為了系統的剖析Greek island的相互作用,作者探討了這些增強子核心序列的作用。在mOSNs的原位Hi-C中, islands H(2kb)、Lipsi(1kb)和Sfaktiria (0.6 kb)的純合子缺失,在包含這些缺失的基因組bins和剩余的Greek islands之間反式相互作用減少,這種效應延伸至較寬的基因組距離(圖2a-c)。值得注意的是,積累的反式Greek island接觸減少與在細胞中觀察到的OR基因的轉錄下調相關圖2c)。小至0.6 kb的DNA在數百個堿基上協調基因組接觸,類似于ZIP元素影響酵母菌或Igk增強子的核定位,從而影響了pre-B細胞中免疫球蛋白位點的定位。三重增強子刪除對集群的影響表明,這些額外的序列參與OR集群之間的交互。
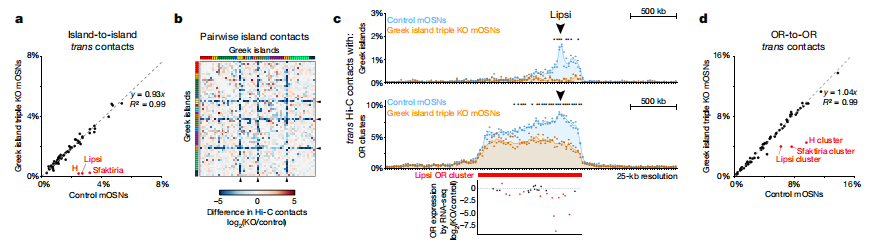
圖2 Greek islands增強子缺失對集群的影響
3. 蛋白質的區域化調節
接下來,作者研究了Greek-island-bound結合轉錄因子在ORs區隔中的作用,在水平基底細胞(HBCs)中刪除LHX2,然后用甲基咪唑誘導這些細胞分化,以TdTomato強度為標記,鑒定了細胞群體,其中最暗的是由HBC-衍生的INPs和mOSNs組成。熒光活化細胞FAC-sorted的RNA測序(RNA-seq)結果顯示,LHX2的早期缺失導致嗅覺神經元(OSN)譜系發育遲緩,INP-特異性標志物增多。考慮到分化缺陷和可能的細胞標識變化,FAC-sorted細胞反式OR和反式 Greek island 接觸明顯減少(圖3a-d)。在早期的LHX2敲除細胞中,染色體間相互作用的頻率仍然很高,但在1000個最強的反式接觸中OR-OR接觸僅占16%。在mOSNs中LHX2的晚期缺失也減少了反式OR接觸,但沒有早期缺失多(圖3a, c)。LHX2的缺失使Greek islands之間的反式接觸和遠距離順式接觸減少(圖3b, d),與OR下調相一致。
為了了解LHX2是如何穩定的與Greek island接觸的,作者研究了LHX2(LIM區域蛋白)是否招募了邊緣結合蛋白(LDB1)。作者對LDB1進行的染色質免疫沉淀和測序(ChIP-seq),發現mOSNs與LHX2峰有密切重疊。與此一致的是,每個Greek island都以LHX2-dependent的方式被LDB1綁定。在mOSNs中刪除LDB1,導致反式Greek island和遠距離順式Greek island相互作用的強烈減少(圖4a, b),OR集群之間反式交互減少,在全基因組中效應更弱。值得注意的是,RNA-seq顯示LDB1的缺失導致ORs廣泛的轉錄下調(圖4c),其可能受限于OR家族(圖4d)。
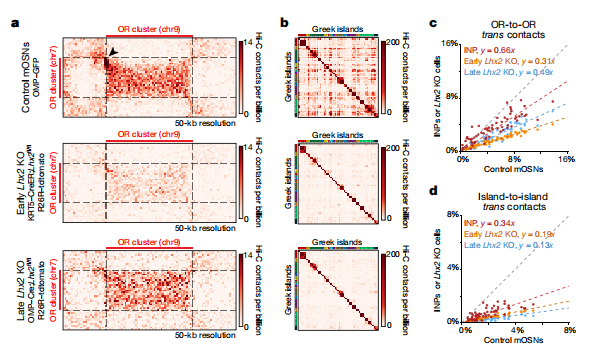
圖3 LHX2是Greek island中心形成OR分隔以及裝配和穩定的基礎
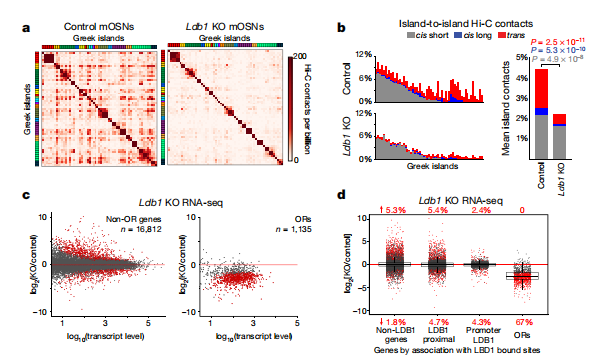
圖4 LDB1是Greek island中心穩定和OR轉錄的基礎
4. Greek island中心與活躍的OR基因有關
為了檢測Greek island中心是否通過與OR基因直接相互作用調節OR轉錄,作者在OR基因 Olfr16、Olfr17和Olfr1507進行了原位Hi-C實驗。在這些OSN種群中,整體OR集群網絡與Greek island的相互作用在很大程度上是相同的,但也觀察到OSN類型特異性變異。然而,在每一種OSN類型中,轉錄活躍的OR始終與Greek islands形成頻繁的相互作用。例如,在OLFR16+ OSNs中,OLFR16位點與Greek islands的順式長交互和反式相互作用強烈,5%的Hi-C接觸點映射到Olfr16。在OLFR17+OSNs和OLFR1507+ OSNs中,它主要與附近的Greek islands相互作用(圖5a, b)。值得注意的是,在OLFR16+細胞中,相對于完整的OR譜,Greek island上的聯系在Olfr16位點得到額外的加強(圖5b, 5c)。因此,通過基因與Greek islands的累積相互作用,原位Hi-C可以準確地從近1000個基因中識別到轉錄活躍的OR。
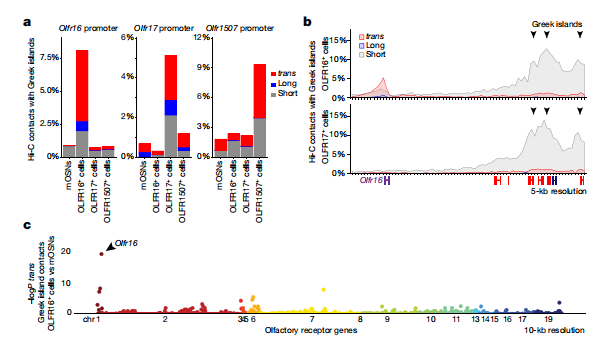
圖5 Greek island中心與OR特異性轉錄活性位點的相互作用
文章小結
本文作者利用原位Hi-C、RNA-seq等技術,發現小鼠嗅覺感受器OR基因間隔以分層的方式形成,順式接觸首先出現,反式作用隨著分化而加強。OR隔間內的Greek islands增強子的缺失對集群之間的交互有影響,轉錄因子LHX2的缺失使Greek islands之間的反式接觸和遠距離順式接觸減少,并發現邊緣結合蛋白LDB1是Greek island中心穩定和OR轉錄的基礎。最后通過原位Hi-C實驗,發現Greek island中心通過與OR基因直接相互作用調節OR轉錄。
解析文獻
Kevin Monahan, Adan Horta, et al. LHX2- and LDB1-mediated trans interactions regulate olfactory receptor choice [J]. Nature, 2019, 565 , 448–453.
相關服務
Hi-C是以整個細胞核為研究對象,利用高通量測序技術,結合生物信息分析方法,研究全基因組范圍內整個染色質DNA在空間位置上的關系,獲得高分辨率的染色質調控元件相互作用圖譜。Hi-C可以與RNA-Seq、ChIP-Seq、ATAC-Seq等數據進行聯合分析,從基因調控網絡和表觀遺傳網絡來闡述生物體性狀形成的相關機制。
武漢金開瑞生物工程有限公司提供的Hi-C技術,是一種新型、高效、經濟的研究方法,幫助您解析基因組三維結構。
最新動態
-
09.23
中藥的現代詮釋:外泌體如何革新傳統醫學?
-
07.02
1+1>2!深度解析RNA測序數據挖掘邏輯和后期實驗設計思路,輕松研獲10+ SCI
-
07.01
“稻”亦有道——盤點近期水稻研究的重大突破
-
06.28
科學與美學的結合體:植物亞細胞定位技術詳解
-
06.28
“聚焦新質生產力,激發科研新動能”|LCA躋身蛋白互作研究的新銳力量
-
06.05
知無不“研”|一文讀懂免疫共沉淀技術(Co-IP)
-
05.14
四大研究利器(Co-IP、BIFC、Y2H、GST pull-down)助力速配蛋白互作“最佳拍檔”
-
05.14
高效、精準、直觀、實時——取經“蛋白互作研究翹楚”BIFC!
-
05.14
轉染效率低、干擾效果差、重復性欠佳...siRNA研究頻遇“攔路虎”怎么辦?
-
04.22
一文讀懂EMSA技術核心要點,讓“emsa” 秒變“easy”
 X
X 






