
多組學聯合揭示糖酵解和KEAP1-NRF2信號轉導的整合機制
信息來源:金開瑞 作者:genecreate 發布時間:2019-12-31 11:31:43
期刊:Nature
IF:41.577
技術:定量蛋白質組、靶向代謝組、PRM
研究背景:
細胞代謝和信號調節整合對于維持細胞穩態是極其必需的。細胞內生、固有活性代謝物能夠調節蛋白質功能,進而影響細胞的生命活動。KEAP1是一種重要的“親電性傳感器”蛋白,可對內源性和外源性分子做出反應,其共價修飾會導致 NRF2 的累積,從而啟動細胞保護基因轉錄。因此,作者希望能揭示糖酵解和NRF2信號傳導之間的直接聯系。
研究結果:
1. IMR32細胞表型篩選,確定激活 NRF2信號傳導的化合物
為了發現KEAP1-NRF2信號傳導途徑的非共價調節因子,以及其潛在調節的新作用機制,作者使用NRF2依賴性熒光素酶報告基因(pTI-ARE-LUC)進行了基于IMR32細胞的高通量表型篩選。從多種雜環化合物庫中,作者鑒定到一種CBR-470-0化合物,其系列化合物(如CBR-470-1)處理IMR32細胞后可導致NRF2蛋白的積累呈現時間依賴效應(圖1b)。根據IMR32細胞(CBR-470-1處理24h后)表達譜顯示,最顯著富集的基因組是由NRF2靶基因組成的“NFE2L2 targets” (圖1c-1d),這表明CBR-470-系列化合物可以在體外誘導NRF2信號通路激活。
接下來,作者進一步確定CBR-470-系列化合物參與體內的誘導過程。Balb/C小鼠口服CBR-470-2化合物,通過qRT-PCR檢測在幾個器官中NRF2靶基因的表達情況,發現在皮膚中觀察到NQO1和HMOX1的轉物水平呈現劑量依賴性增加(圖1e)。已有研究表明NRF2可以有效防止紫外線照射導致的光老化表型和皮膚癌變,進而作者在急性UV損傷小鼠模型中評估CBR-470-2的活性,發現CBR-470-2可以減少紫外線照射后的小鼠模型表皮厚度,與NRF2細胞保護程序的激活一致(圖1g-1h);由此表明CBR-470-系列化合物可以在體內誘導NRF2信號通路激活。
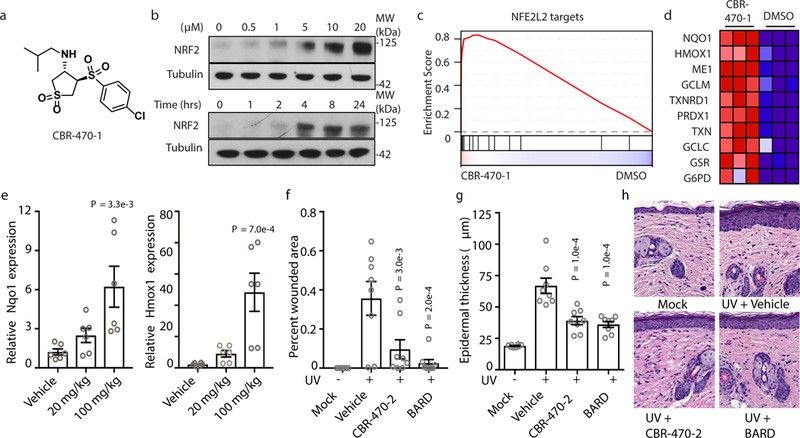
圖1 CBR-470-系列化合物在體外和體內均激活NRF2信號傳導
2. CBR-470-1激活NRF2信號傳導的機制
為了確定CBR-470-1激活NRF2信號傳導的機制,作者合成了含有生物素和二氮雜萘取代基的熒光親和探針(稱為CBR-470-PAP),并使用5μM劑量去處理IMR32細胞1小時,在紫外線照射后進行細胞裂解液的抗生物素蛋白質印跡分析(圖2a);再接著通過生化分離和LC-MS/MS分析磷酸甘油酸激酶1(PGK1)是否作為CBR-470-PAP的潛在靶標(圖2b-2c)。體外重組蛋白的結合實驗表明CBR-470-PAP選擇性標記了PGK1;而IMR32細胞中PGK1蛋白的敲低和過表達實驗發現,CBR-470-1處理組的 EC50值分別表現出降低和增加(圖2d-2e),且烯醇酶1(PGK1下游的酶)消耗加快;靶向代謝物組學分析發現,CBR-470-1處理的IMR32細胞中PGK1 上游代謝物水平快速增加以及下游代謝物快速消耗;上述結果表明,糖酵解中間體可參與NRF2信號傳導,CBR-470-1可通過調節PGK1活性來影響糖酵解途徑和NRF2活化過程。
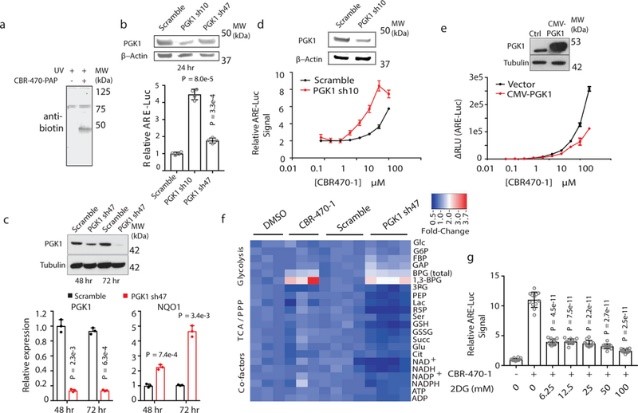
圖2 糖酵解依賴 CBR-470-1 激活 NRF2 信號傳導
3. MGx介導了糖酵解和KEAP1-NRF2信號通路的直接聯系
作者先研究由PGK1直接代謝的1,3-BPG是否可能通過由KEAP1的磷酸甘油基-賴氨酸(PGK)修飾來參與KEAP1-NRF2途徑的信號傳導。在CBR-470-1處理IMR32細胞30分鐘后,1,3-BPG水平升高,KEAP1水平沒有明顯改變;但是WB結果顯示CBR-470-1發生作用依賴于高分子量 KEAP1(HMM-KEAP1)的形成,其分子量大約是單體 KEAP1 的兩倍(圖3a)。除1,3-BPG外,其他糖酵解的中心代謝產物,如磷酸丙糖異構體D-甘油醛-3-磷酸(GAP)和磷酸二羥丙酮(DHAP),以及它們的非酶解產物甲基乙二醛(MGx),這些親電羰基化合物都可以作為反應官能團修飾蛋白殘基。在這些候選物中,僅有MGx處理的細胞裂解物或活細胞可導致HMW-KEAP1的選擇性形成(圖3b-3c),并功能性地激活下游NRF2靶基因NQO1和HMOX1的表達(圖3d);同時衍生化MGx的靶向LC-MS檢測也證實CBR-470-1處理細胞的最初幾小時內MGx水平顯著地升高(圖3e)。因此上述結果說明糖酵解和由MGx直接修飾KEAP1介導的KEAP1-NRF2信號通路之間存在直接聯系。
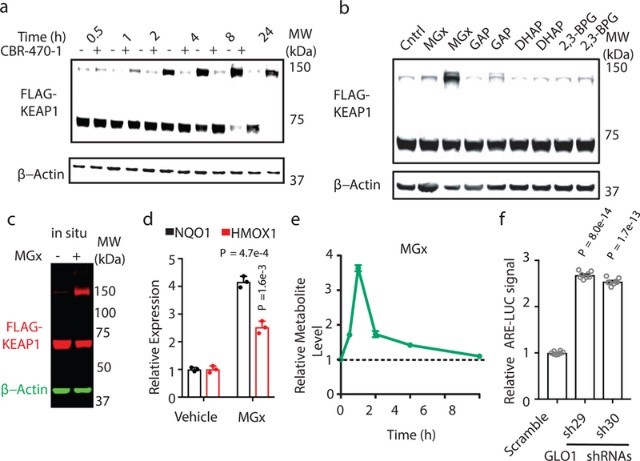
圖3 MGx修飾KEAP1形成共價的高分子量二聚體并激活NRF2信號傳導
4. SILAC 定量蛋白質組學和PRM靶向蛋白質組學確定了影響 HMM-KEAP1 形成的關鍵結構域
接下來,作者進一步選用SILAC 定量蛋白質組學方法確定HMW的結構域和殘基(圖4a),結果表明NTR(N-末端區域,氨基酸1-50)和BTB結構域(氨基酸150-169)作為可能參與HMW的候選結構域和殘基,它們響應了CBR-470-1誘導的HMW-KEAP1二聚體形成過程(圖4b-4d)。同時,作者檢測了這些結構域內的十多個C-to-S,K-to-M/R和R-to-A突變,以及KEAP1中其他已知的功能殘基對HMW-KEAP1形成的影響;結果發現兩個精氨酸殘基突變(NTR結構域的R15和BTB結構域的R135)顯著但不完全地減少了HMW-KEAP1的形成,而BTB結構域中C151S突變對HMW-KEAP1形成的幾乎完全抑制(圖5a)。由此,作者通過合成一個含有半胱氨酸和精氨酸的模型肽,來確定MGx可能介導上述殘基之間的修飾;采用甘氨酸接頭分開模型肽,并在生理溫度和pH下用MGx處理過夜,可發現形成翻譯后修飾出新的甲基咪唑(MICA)。
隨后,作者在純化MICA后通過一系列一維和二維NMR實驗證實了其結構(圖5d);并采用CBR-470-1或MGx處理細胞,經過凝膠分離HMW-KEAP1和單體KEAP1后進行LC-MS/MS分析,來確定是否在KEAP1蛋白內發生MICA修飾(圖5c-5d)。在來自CBR-470-1和MGx處理的分離的HMW-KEAP1中鑒定了在C151和R135之間具有MICA交聯肽,但在分離的單體KEAP1中未鑒定到(圖5e)。此外,PRM的并行反應監測也證實了HMW-KEAP1中獨特存在的十幾個母-子離子躍遷的存在和共洗脫的存在(圖5f;圖6a-6b)。綜上所述,作者發現糖酵解代謝過程是通過反應性糖酵解代謝物MGx與前哨蛋白KEAP1的直接相互作用來偶聯NRF2依賴性基因的表達,并形成穩定且新型的翻譯后修飾蛋白質來介導糖酵解途徑(圖5g)。
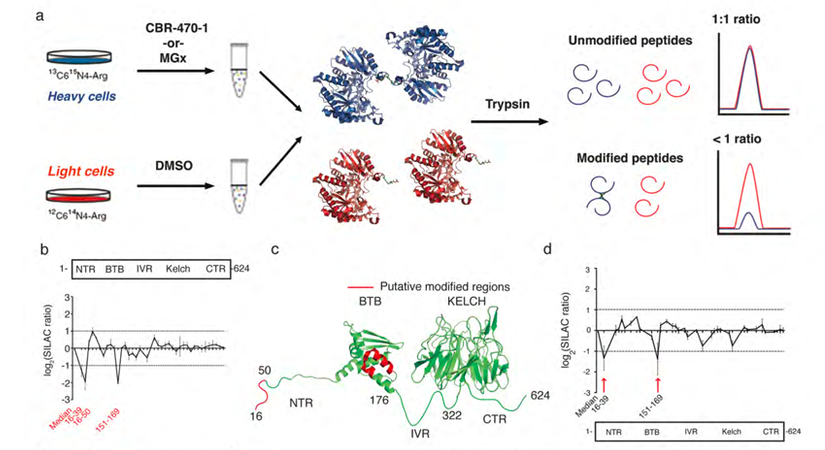
圖4 基于SILAC的KEAP1修飾蛋白質組學表征HMM-KEAP1結構域
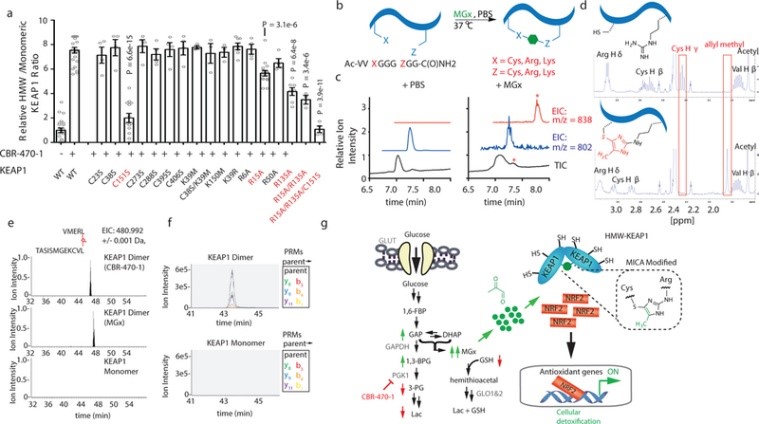
圖5 甲基乙二醛在KEAP1中的近端半胱氨酸和精氨酸殘基之間形成新的翻譯后修飾
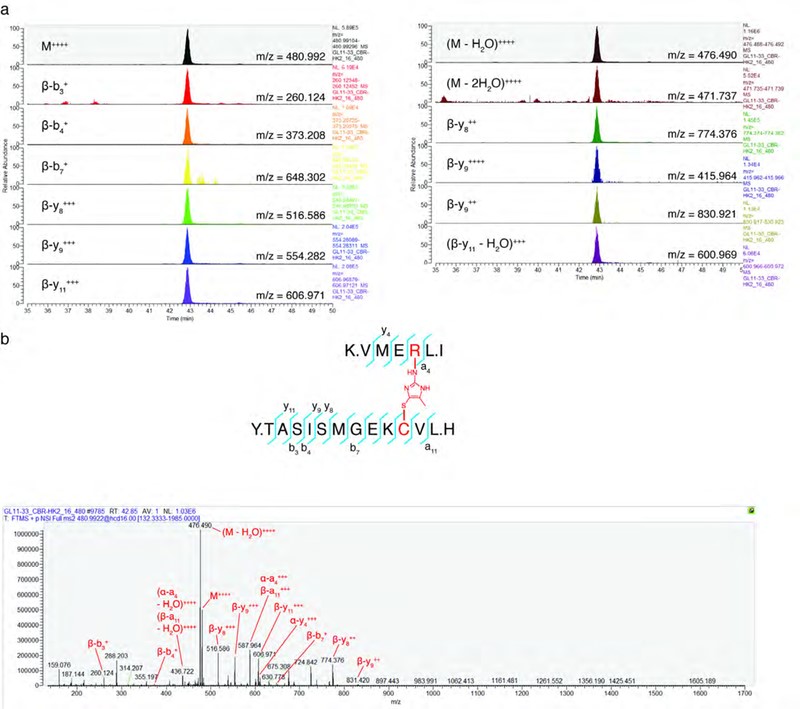
圖6 PRM分析MICA交聯肽
研究結論:
作者確定了糖酵解酶PGK1的小分子抑制劑,并揭示了糖酵解和NRF2信號傳導之間的直接聯系。PGK1的抑制導致活性代謝物MGx的積累,其選擇性地修飾KEAP1以便在近端半胱氨酸和精氨酸殘基(MICA)之間形成甲基咪唑交聯;該翻譯后修飾的過程導致KEAP1的二聚化、NRF2的積累和NRF2轉錄程序的激活。總的來說,在糖酵解和KEAP1-NRF2信號傳導之間存在著直接的通信聯系,由此更加深入地了解細胞應激反應后的代謝調控機制。
金開瑞推薦技術服務:
新技術的不斷涌現,加快了多組學研究向定量化、高通量的發展,在分子水平研究基因組學、轉錄組學、蛋白質組學和代謝組學,已成為人們發現生命化學物質基礎和深入了解其分子機制的新方向。
多組學數據整合分析,有利于系統性地研究臨床發病機理、確認疾病靶點,發現生物標志物與進行疾病早期診斷,從而對個體化治療和用藥指導發揮重要作用。
金開瑞現提供下述多組學技術服務,并贈送對應關聯分析:
| 多組學項目 | 贈送關聯分析類型 |
| 轉錄組+蛋白質組 |
轉錄組+蛋白質組定性分析
轉錄組+蛋白質組定量關聯分析
轉錄組+蛋白質組層次聚類分析
轉錄組+蛋白質組GO、Pathway富集分析 |
| 蛋白質組+代謝組 |
蛋白質組+代謝組 PCA 比較分析
差異蛋白+差異代謝物表達量相關性分析
差異蛋白+差異代謝物Pathway分析
蛋白質組+代謝組相關性分析(矩陣熱圖和相關性網絡)
加權基因共表達網絡分析(WGCNA,≥15 sample)
|
| 轉錄組+蛋白質組+代謝組 |
表達量相關性分析
差異表達分析
Pathway富集分析
|
活動進行中~ Label-free修飾引物
最新動態
-
09.23
中藥的現代詮釋:外泌體如何革新傳統醫學?
-
07.02
1+1>2!深度解析RNA測序數據挖掘邏輯和后期實驗設計思路,輕松研獲10+ SCI
-
07.01
“稻”亦有道——盤點近期水稻研究的重大突破
-
06.28
科學與美學的結合體:植物亞細胞定位技術詳解
-
06.28
“聚焦新質生產力,激發科研新動能”|LCA躋身蛋白互作研究的新銳力量
-
06.05
知無不“研”|一文讀懂免疫共沉淀技術(Co-IP)
-
05.14
四大研究利器(Co-IP、BIFC、Y2H、GST pull-down)助力速配蛋白互作“最佳拍檔”
-
05.14
高效、精準、直觀、實時——取經“蛋白互作研究翹楚”BIFC!
-
05.14
轉染效率低、干擾效果差、重復性欠佳...siRNA研究頻遇“攔路虎”怎么辦?
-
04.22
一文讀懂EMSA技術核心要點,讓“emsa” 秒變“easy”
 X
X 






